

Actin bodies in yeast quiescent cells: an immediately available actin reserve?
- PMID: 16914523
- PMCID: PMC1635378
- DOI: 10.1091/mbc.e06-04-0282
Actin bodies in yeast quiescent cells: an immediately available actin reserve?
Abstract
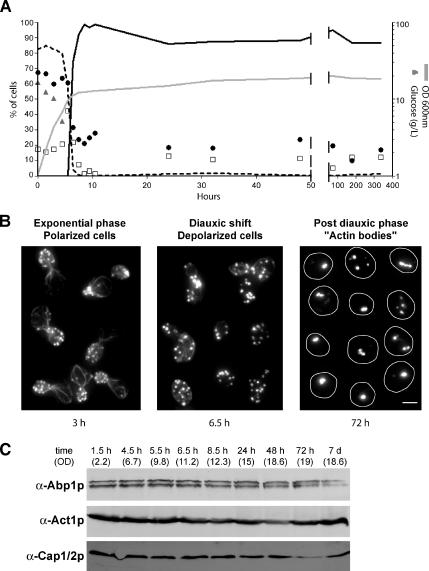
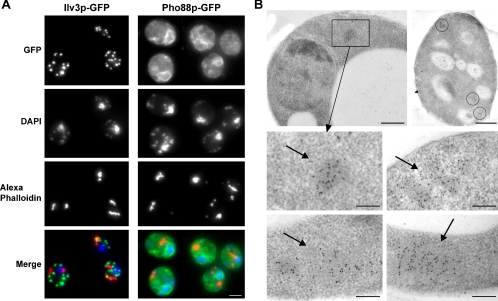
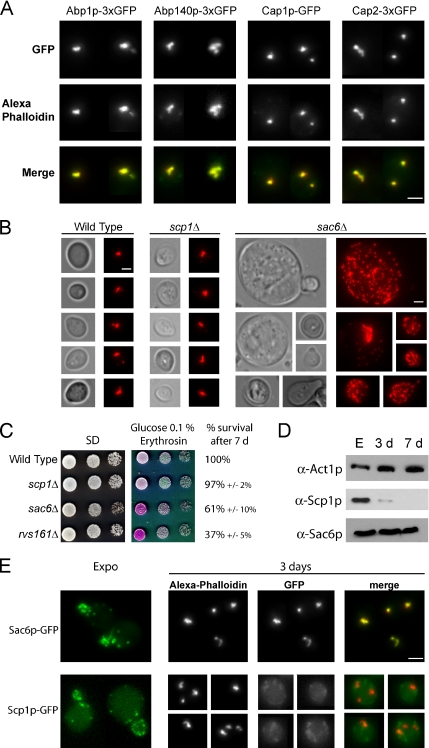
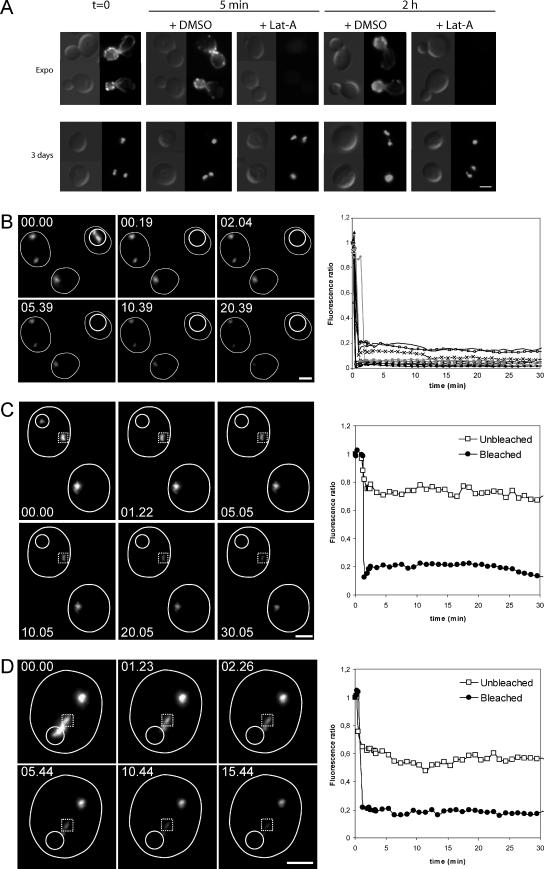
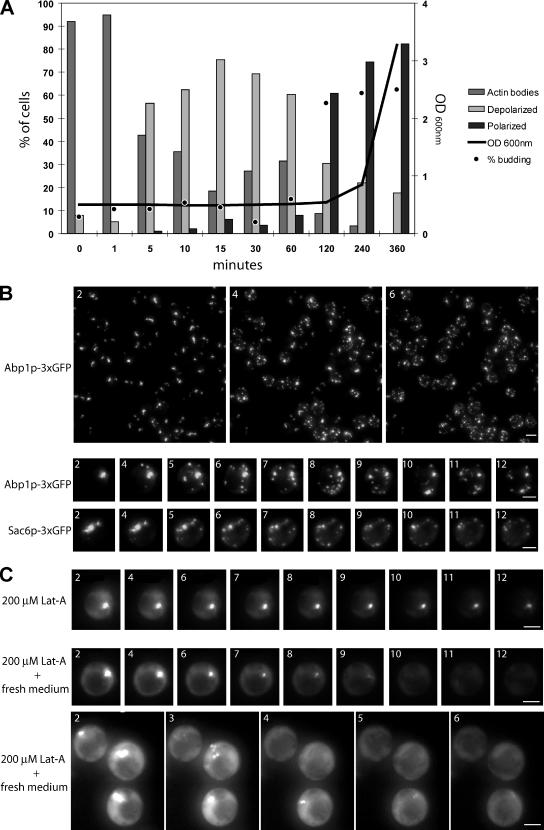
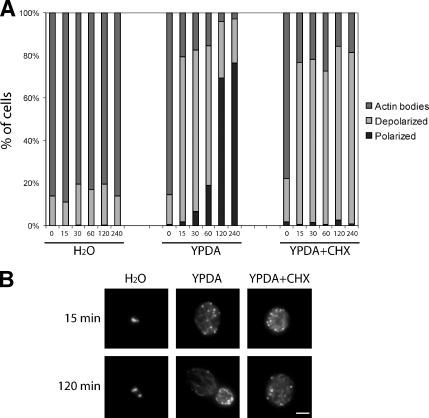
Most eukaryotic cells spend most of their life in a quiescent state, poised to respond to specific signals to proliferate. In Saccharomyces cerevisiae, entry into and exit from quiescence are dependent only on the availability of nutrients in the environment. The transition from quiescence to proliferation requires not only drastic metabolic changes but also a complete remodeling of various cellular structures. Here, we describe an actin cytoskeleton organization specific of the yeast quiescent state. When cells cease to divide, actin is reorganized into structures that we named "actin bodies." We show that actin bodies contain F-actin and several actin-binding proteins such as fimbrin and capping protein. Furthermore, by contrast to actin patches or cables, actin bodies are mostly immobile, and we could not detect any actin filament turnover. Finally, we show that upon cells refeeding, actin bodies rapidly disappear and actin cables and patches can be assembled in the absence of de novo protein synthesis. This led us to propose that actin bodies are a reserve of actin that can be immediately mobilized for actin cables and patches formation upon reentry into a proliferation cycle.
Figures
Similar articles
-
Interactions among a fimbrin, a capping protein, and an actin-depolymerizing factor in organization of the fission yeast actin cytoskeleton.Mol Biol Cell. 2001 Nov;12(11):3515-26. doi: 10.1091/mbc.12.11.3515. Mol Biol Cell. 2001. PMID: 11694585 Free PMC article.
-
Yeast Aim21/Tda2 both regulates free actin by reducing barbed end assembly and forms a complex with Cap1/Cap2 to balance actin assembly between patches and cables.Mol Biol Cell. 2018 Apr 15;29(8):923-936. doi: 10.1091/mbc.E17-10-0592. Epub 2018 Mar 30. Mol Biol Cell. 2018. PMID: 29467252 Free PMC article.
-
The yeast actin cytoskeleton: from cellular function to biochemical mechanism.Microbiol Mol Biol Rev. 2006 Sep;70(3):605-45. doi: 10.1128/MMBR.00013-06. Microbiol Mol Biol Rev. 2006. PMID: 16959963 Free PMC article. Review.
-
The Stationary-Phase Cells of Saccharomyces cerevisiae Display Dynamic Actin Filaments Required for Processes Extending Chronological Life Span.Mol Cell Biol. 2015 Nov;35(22):3892-908. doi: 10.1128/MCB.00811-15. Epub 2015 Sep 8. Mol Cell Biol. 2015. PMID: 26351139 Free PMC article.
-
Control of reorganization of the actin cytoskeleton by Rho family small GTP-binding proteins in yeast.Curr Opin Cell Biol. 1998 Feb;10(1):112-6. doi: 10.1016/s0955-0674(98)80093-8. Curr Opin Cell Biol. 1998. PMID: 9484602 Review.
Cited by
-
Integrity of the Saccharomyces cerevisiae Rpn11 protein is critical for formation of proteasome storage granules (PSG) and survival in stationary phase.PLoS One. 2013 Aug 6;8(8):e70357. doi: 10.1371/journal.pone.0070357. Print 2013. PLoS One. 2013. PMID: 23936414 Free PMC article.
-
Proteins implicated in mediating self-incompatibility-induced alterations to the actin cytoskeleton of Papaver pollen.Ann Bot. 2011 Sep;108(4):659-75. doi: 10.1093/aob/mcr022. Epub 2011 Feb 13. Ann Bot. 2011. PMID: 21320881 Free PMC article. Review.
-
A stable microtubule bundle formed through an orchestrated multistep process controls quiescence exit.Elife. 2024 Mar 25;12:RP89958. doi: 10.7554/eLife.89958. Elife. 2024. PMID: 38527106 Free PMC article.
-
Self-incompatibility-induced programmed cell death in field poppy pollen involves dramatic acidification of the incompatible pollen tube cytosol.Plant Physiol. 2015 Mar;167(3):766-79. doi: 10.1104/pp.114.252742. Epub 2015 Jan 28. Plant Physiol. 2015. PMID: 25630437 Free PMC article.
-
Nutritional control of growth and development in yeast.Genetics. 2012 Sep;192(1):73-105. doi: 10.1534/genetics.111.135731. Genetics. 2012. PMID: 22964838 Free PMC article. Review.
References
-
- Adams A. E., Botstein D., Drubin D. G. Requirement of yeast fimbrin for actin organization and morphogenesis in vivo. Nature. 1991;354:404–408. - PubMed
-
- Asakura T. Isolation and characterization of a novel actin filament-binding protein from Saccharomyces cerevisiae. Oncogene. 1998;16:121–130. - PubMed
-
- Binley K. M., Radcliffe P. A., Trevethick J., Duffy K. A., Sudbery P. E. The yeast PRS3 gene is required for cell integrity, cell cycle arrest upon nutrient deprivation, ion homeostasis and the proper organization of the actin cytoskeleton. Yeast. 1999;15:1459–1469. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
