

Scaffolding protein INAD regulates deactivation of vision by promoting phosphorylation of transient receptor potential by eye protein kinase C in Drosophila
- PMID: 16914683
- PMCID: PMC1577681
- DOI: 10.1523/JNEUROSCI.1478-06.2006
Scaffolding protein INAD regulates deactivation of vision by promoting phosphorylation of transient receptor potential by eye protein kinase C in Drosophila
Abstract
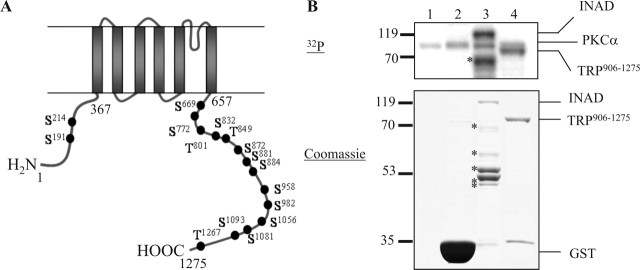
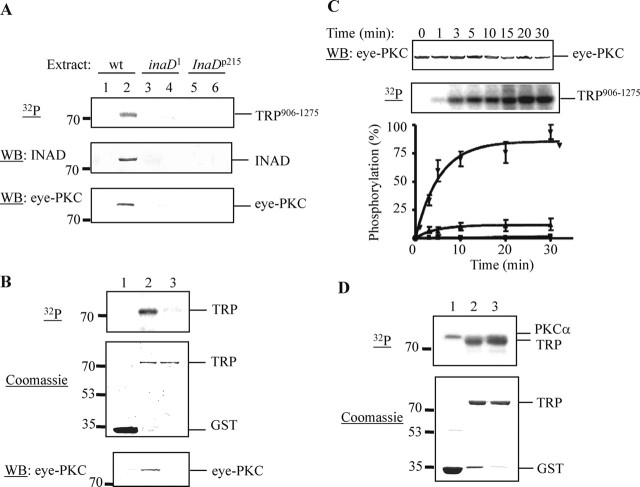
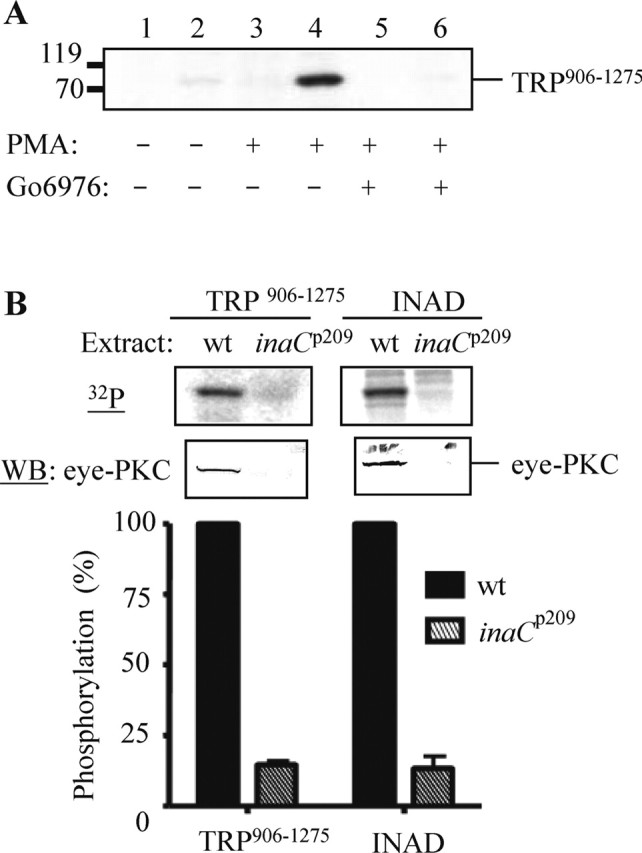
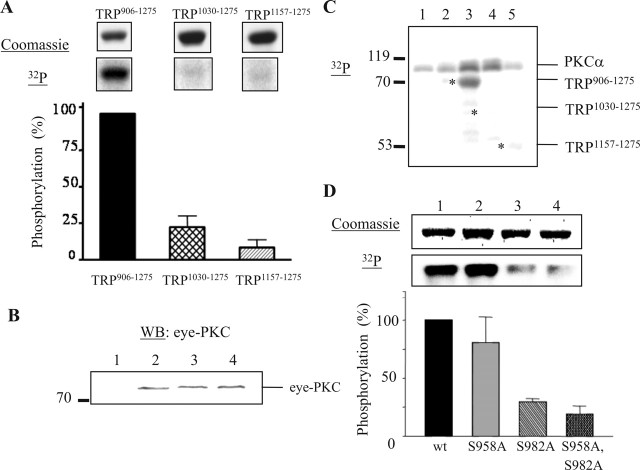
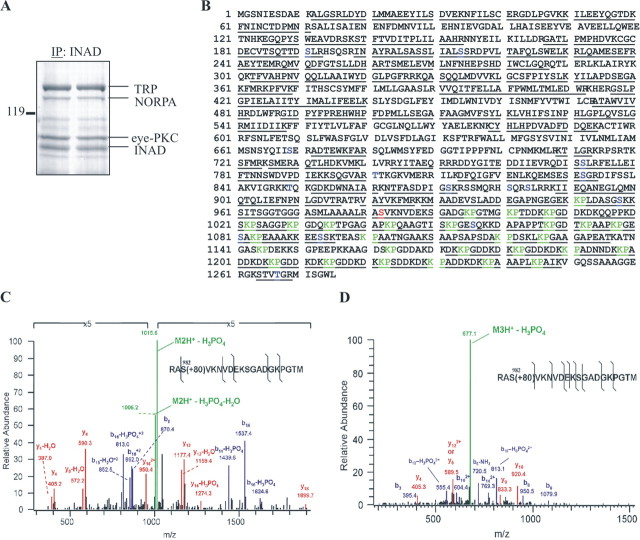
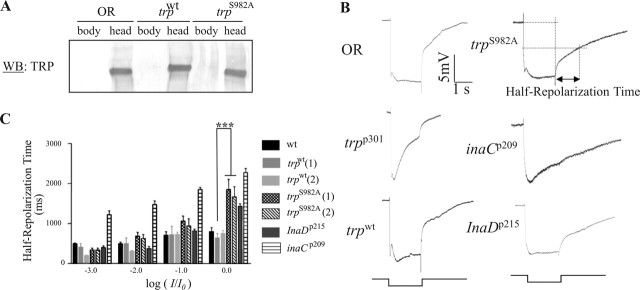
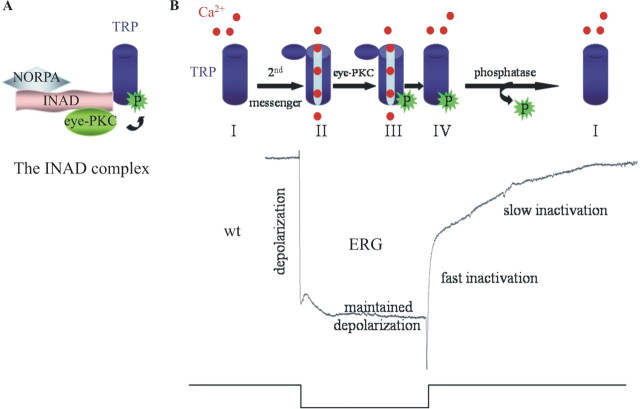
Drosophila visual signaling is one of the fastest G-protein-coupled transduction cascades, because effector and modulatory proteins are organized into a macromolecular complex ("transducisome"). Assembly of the complex is orchestrated by inactivation no afterpotential D (INAD), which colocalizes the transient receptor potential (TRP) Ca2+ channel, phospholipase Cbeta, and eye protein kinase C (eye-PKC), for more efficient signal transduction. Eye-PKC is critical for deactivation of vision. Moreover, deactivation is regulated by the interaction between INAD and TRP, because abrogation of this interaction in InaD(p215) results in slow deactivation similar to that of inaC(p209) lacking eye-PKC. To elucidate the mechanisms whereby eye-PKC modulates deactivation, here we demonstrate that eye-PKC, via tethering to INAD, phosphorylates TRP in vitro. We reveal that Ser982 of TRP is phosphorylated by eye-PKC in vitro and, importantly, in the fly eye, as shown by mass spectrometry. Furthermore, transgenic expression of modified TRP bearing an Ala substitution leads to slow deactivation of the visual response similar to that of InaD(p215). These results suggest that the INAD macromolecular complex plays an essential role in termination of the light response by promoting efficient phosphorylation at Ser982 of TRP for fast deactivation of the visual signaling.
Figures
References
-
- Adamski FM, Zhu MY, Bahiraei F, Shieh BH (1998). Interaction of eye protein kinase C and INAD in Drosophila. Localization of binding domains and electrophysiological characterization of a loss of association in transgenic flies. J Biol Chem 273:17713–17719. - PubMed
-
- Ahmmed GU, Mehta D, Vogel S, Holinstat M, Paria BC, Tiruppathi C, Malik AB (2004). Protein kinase Cα phosphorylates the TRPC1 channel and regulates store-operated Ca2+ entry in endothelial cells. J Biol Chem 279:20941–20949. - PubMed
-
- Bloomquist BT, Shortridge RD, Schneuwly S, Perdew M, Montell C, Steller H, Rubin G, Pak WL (1988). Isolation of a putative phospholipase C gene of Drosophila, norpA, and its role in phototransduction. Cell 54:723–733. - PubMed
-
- Chevesich J, Kreuz AJ, Montell C (1997). Requirement for the PDZ domain protein, INAD, for localization of the TRP store-operated channel to a signaling complex. Neuron 18:95–105. - PubMed
-
- Chyb S, Raghu P, Hardie RC (1999). Polyunsaturated fatty acids activate the Drosophila light-sensitive channels TRP and TRPL. Nature 397:255–259. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous