

Binding of SH2-B family members within a potential negative regulatory region maintains JAK2 in an active state
- PMID: 16914724
- PMCID: PMC1592834
- DOI: 10.1128/MCB.00570-06
Binding of SH2-B family members within a potential negative regulatory region maintains JAK2 in an active state
Abstract
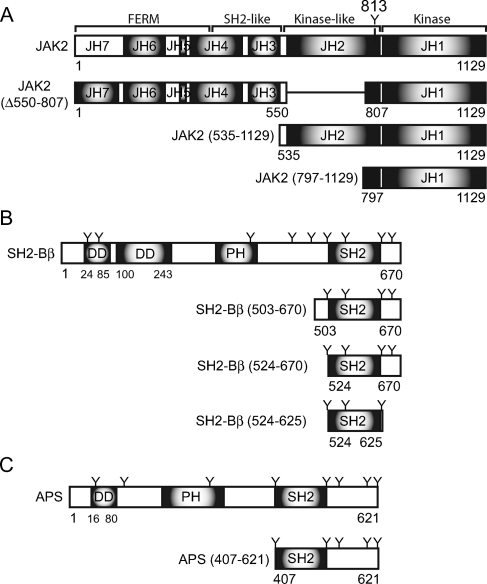
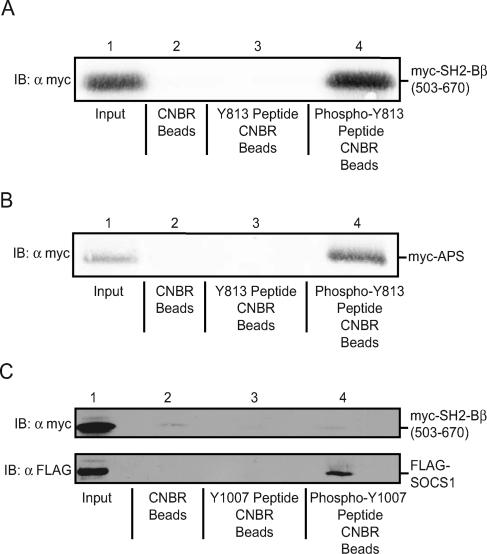
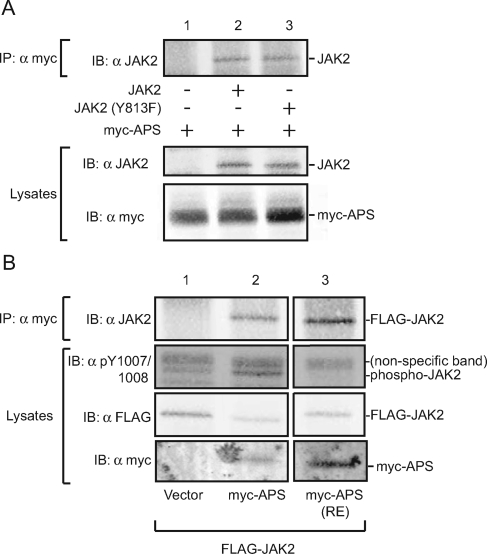
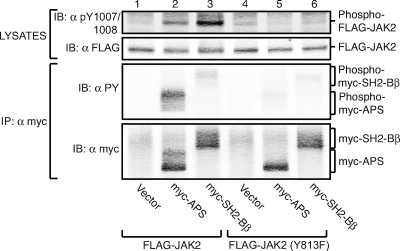
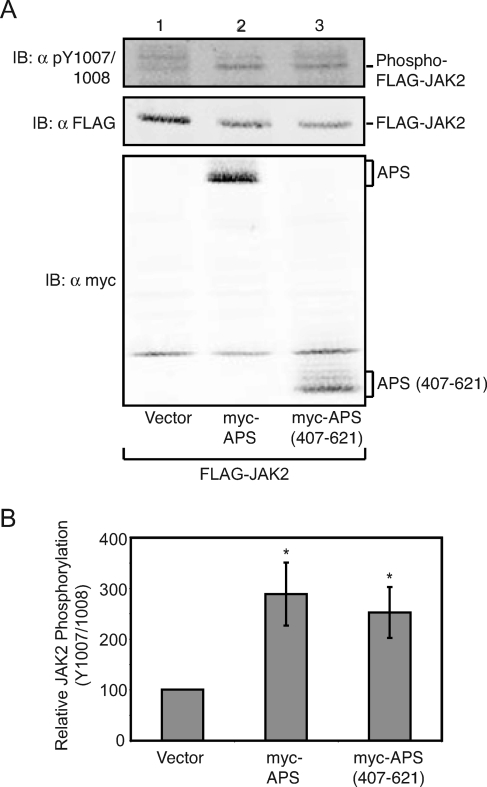
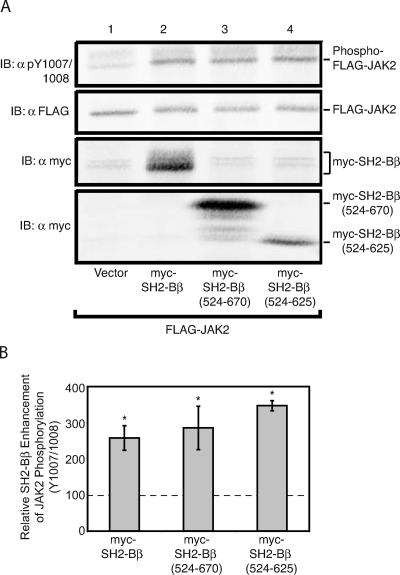
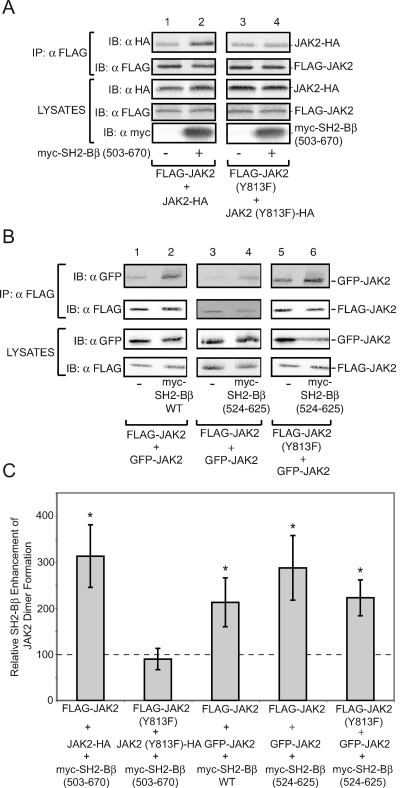
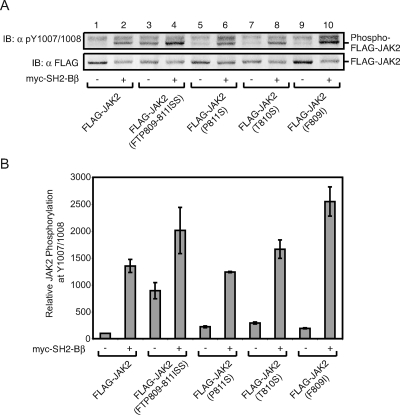
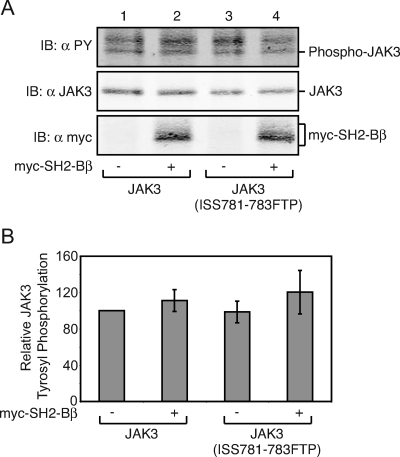
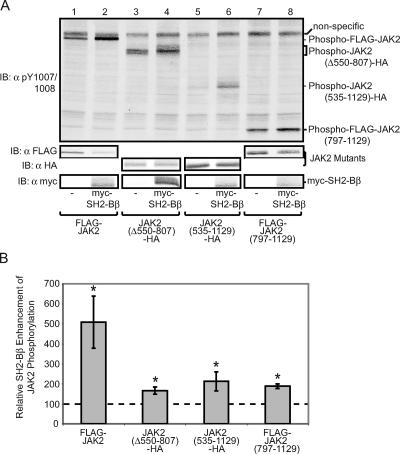
The tyrosine kinase Janus kinase 2 (JAK2) transduces signaling for the majority of known cytokine receptor family members and is constitutively activated in some cancers. Here we examine the mechanisms by which the adapter proteins SH2-Bbeta and APS regulate the activity of JAK2. We show that like SH2-Bbeta, APS binds JAK2 at multiple sites and that binding to phosphotyrosine 813 is essential for APS to increase active JAK2 and to be phosphorylated by JAK2. Binding of APS to a phosphotyrosine 813-independent site inhibits JAK2. Both APS and SH2-Bbeta increase JAK2 activity independent of their N-terminal dimerization domains. SH2-Bbeta-induced increases in JAK2 dimerization require only the SH2 domain and only one SH2-Bbeta to be bound to a JAK2 dimer. JAK2 mutations and truncations revealed that amino acids 809 to 811 in JAK2 are a critical component of a larger regulatory region within JAK2, most likely including amino acids within the JAK homology 1 (JH1) and JH2 domains and possibly the FERM domain. Together, our data suggest that SH2-Bbeta and APS do not activate JAK2 as a consequence of their own dimerization, recruitment of an activator of JAK2, or direct competition with a JAK2 inhibitor for binding to JAK2. Rather, they most likely induce or stabilize an active conformation of JAK2.
Figures
References
-
- Brown, R. J., J. J. Adams, R. A. Pelekanos, Y. Wan, W. J. McKinstry, K. Palethorpe, R. M. Seeber, T. A. Monks, K. A. Eidne, M. W. Parker, and M. J. Waters. 2005. Model for growth hormone receptor activation based on subunit rotation within a receptor dimer. Nat. Struct. Mol. Biol. 12:814-821. - PubMed
-
- Darnell, J. E., Jr., I. M. Kerr, and G. R. Stark. 1994. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 264:1415-1421. - PubMed
-
- Dhe-Paganon, S., E. D. Werner, M. Nishi, L. Hansen, Y. I. Chi, and S. E. Shoelson. 2004. A phenylalanine zipper mediates APS dimerization. Nat. Struct. Mol. Biol. 11:968-974. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK054222/DK/NIDDK NIH HHS/United States
- DK34171/DK/NIDDK NIH HHS/United States
- P60-DK20572/DK/NIDDK NIH HHS/United States
- P30 CA046592/CA/NCI NIH HHS/United States
- R37 DK034171/DK/NIDDK NIH HHS/United States
- P30 CA46592/CA/NCI NIH HHS/United States
- R56 DK054222/DK/NIDDK NIH HHS/United States
- P60-AR20557/AR/NIAMS NIH HHS/United States
- DK54222/DK/NIDDK NIH HHS/United States
- T32-GM08322/GM/NIGMS NIH HHS/United States
- T32 GM007863/GM/NIGMS NIH HHS/United States
- T32 GM008322/GM/NIGMS NIH HHS/United States
- P60 DK020572/DK/NIDDK NIH HHS/United States
- T32 GM07863/GM/NIGMS NIH HHS/United States
- R01 DK034171/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous