

Actin-induced hyperactivation of the Ras signaling pathway leads to apoptosis in Saccharomyces cerevisiae
- PMID: 16914733
- PMCID: PMC1592845
- DOI: 10.1128/MCB.00117-06
Actin-induced hyperactivation of the Ras signaling pathway leads to apoptosis in Saccharomyces cerevisiae
Abstract
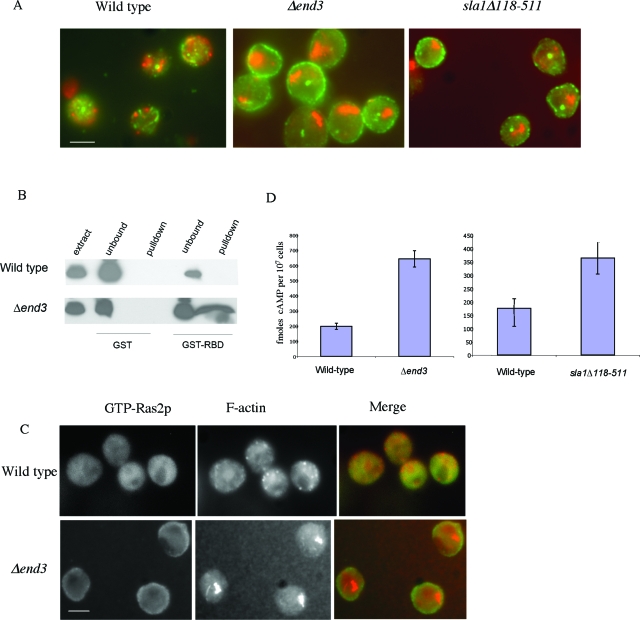
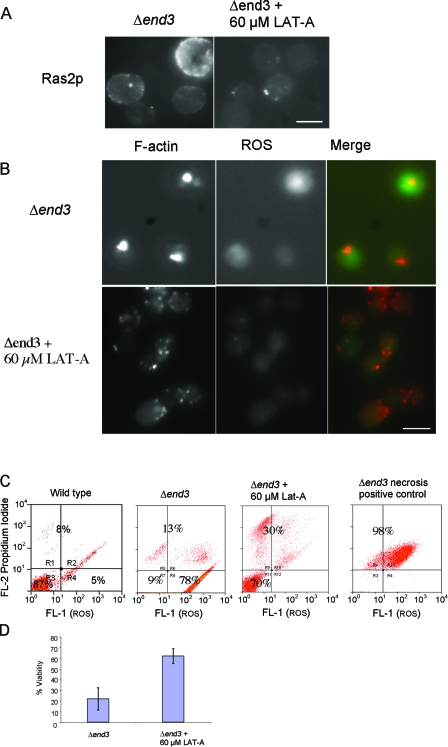
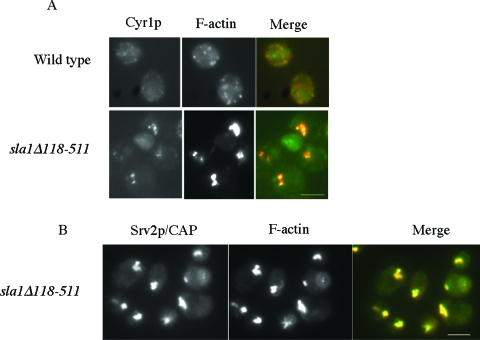
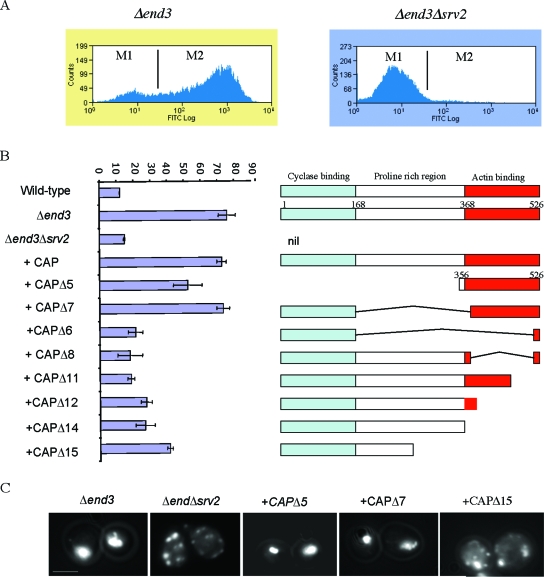
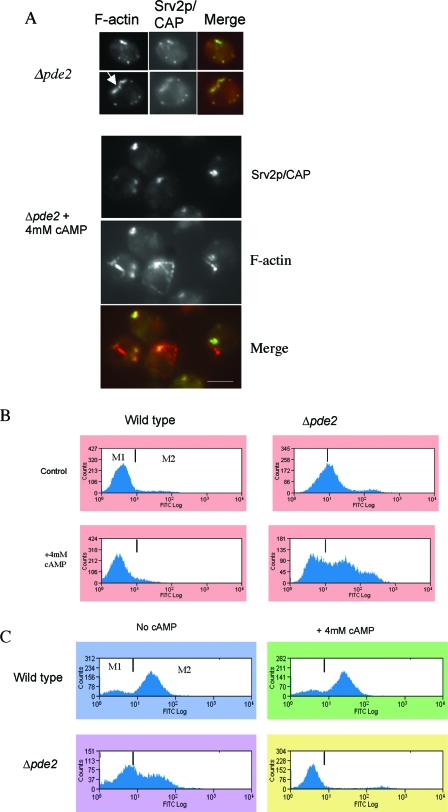
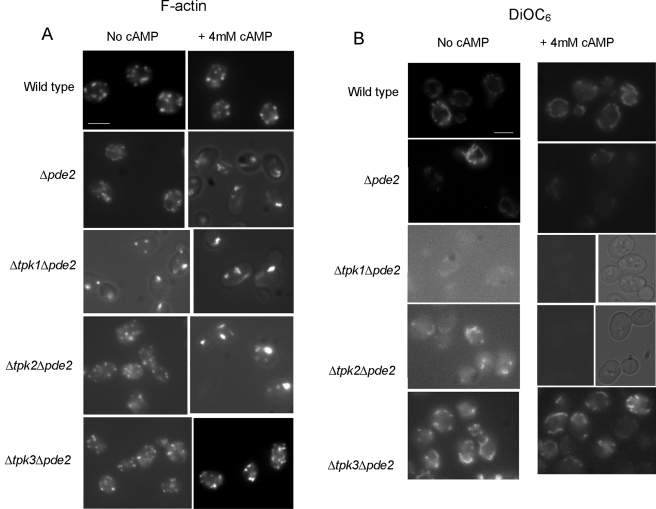
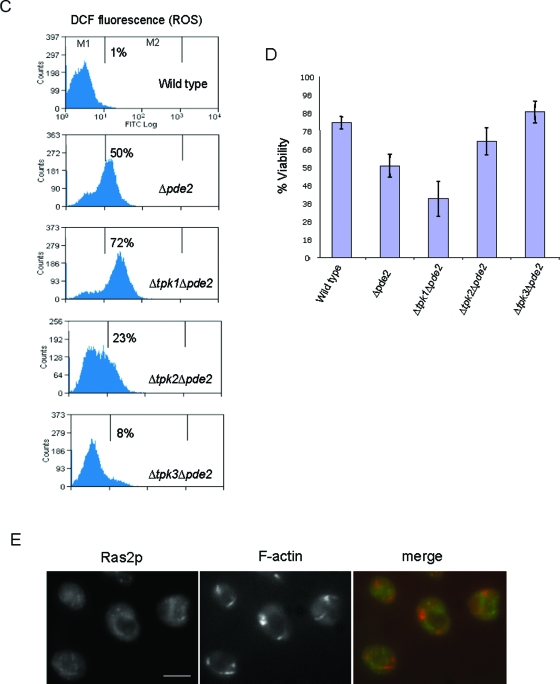
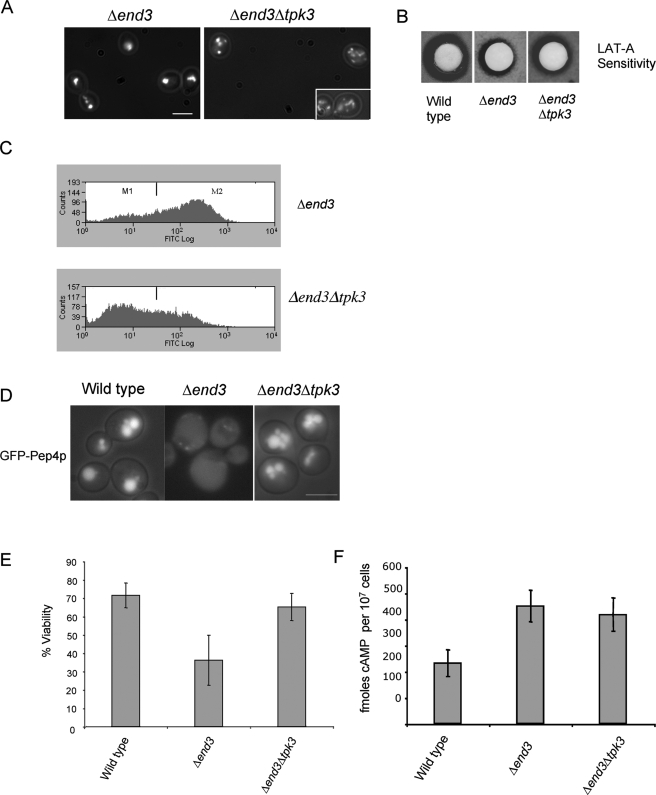
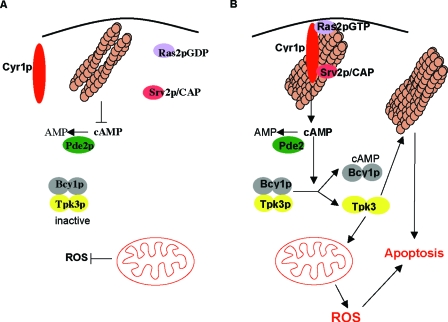
Recent research has revealed a conserved role for the actin cytoskeleton in the regulation of aging and apoptosis among eukaryotes. Here we show that the stabilization of the actin cytoskeleton caused by deletion of Sla1p or End3p leads to hyperactivation of the Ras signaling pathway. The consequent rise in cyclic AMP (cAMP) levels leads to the loss of mitochondrial membrane potential, accumulation of reactive oxygen species (ROS), and cell death. We have established a mechanistic link between Ras signaling and actin by demonstrating that ROS production in actin-stabilized cells is dependent on the G-actin binding region of the cyclase-associated protein Srv2p/CAP. Furthermore, the artificial elevation of cAMP directly mimics the apoptotic phenotypes displayed by actin-stabilized cells. The effect of cAMP elevation in inducing actin-mediated apoptosis functions primarily through the Tpk3p subunit of protein kinase A. This pathway represents the first defined link between environmental sensing, actin remodeling, and apoptosis in Saccharomyces cerevisiae.
Figures
Similar articles
-
Whi2p links nutritional sensing to actin-dependent Ras-cAMP-PKA regulation and apoptosis in yeast.J Cell Sci. 2009 Mar 1;122(Pt 5):706-15. doi: 10.1242/jcs.042424. Epub 2009 Feb 10. J Cell Sci. 2009. PMID: 19208759 Free PMC article.
-
A cytoskeletal localizing domain in the cyclase-associated protein, CAP/Srv2p, regulates access to a distant SH3-binding site.J Biol Chem. 1999 Jul 9;274(28):19985-91. doi: 10.1074/jbc.274.28.19985. J Biol Chem. 1999. PMID: 10391948
-
The actin cytoskeleton, RAS-cAMP signaling and mitochondrial ROS in yeast apoptosis.Trends Cell Biol. 2005 Dec;15(12):637-9. doi: 10.1016/j.tcb.2005.09.011. Epub 2005 Oct 14. Trends Cell Biol. 2005. PMID: 16226888
-
Novel sensing mechanisms and targets for the cAMP-protein kinase A pathway in the yeast Saccharomyces cerevisiae.Mol Microbiol. 1999 Sep;33(5):904-18. doi: 10.1046/j.1365-2958.1999.01538.x. Mol Microbiol. 1999. PMID: 10476026 Review.
-
A role for actin in aging and apoptosis.Biochem Soc Trans. 2005 Dec;33(Pt 6):1260-4. doi: 10.1042/BST0331260. Biochem Soc Trans. 2005. PMID: 16246093 Review.
Cited by
-
Fungal communication requires the MAK-2 pathway elements STE-20 and RAS-2, the NRC-1 adapter STE-50 and the MAP kinase scaffold HAM-5.PLoS Genet. 2014 Nov 20;10(11):e1004762. doi: 10.1371/journal.pgen.1004762. eCollection 2014 Nov. PLoS Genet. 2014. PMID: 25411845 Free PMC article.
-
Oxidative stress and programmed cell death in yeast.Front Oncol. 2012 Jun 20;2:64. doi: 10.3389/fonc.2012.00064. eCollection 2012. Front Oncol. 2012. PMID: 22737670 Free PMC article.
-
Acetic acid treatment in S. cerevisiae creates significant energy deficiency and nutrient starvation that is dependent on the activity of the mitochondrial transcriptional complex Hap2-3-4-5.Front Oncol. 2012 Sep 21;2:118. doi: 10.3389/fonc.2012.00118. eCollection 2012. Front Oncol. 2012. PMID: 23050242 Free PMC article.
-
Yeast cell death pathway requiring AP-3 vesicle trafficking leads to vacuole/lysosome membrane permeabilization.Cell Rep. 2022 Apr 12;39(2):110647. doi: 10.1016/j.celrep.2022.110647. Cell Rep. 2022. PMID: 35417721 Free PMC article.
-
Amphotericin B membrane action: role for two types of ion channels in eliciting cell survival and lethal effects.J Membr Biol. 2010 Dec;238(1-3):1-20. doi: 10.1007/s00232-010-9313-y. Epub 2010 Nov 18. J Membr Biol. 2010. PMID: 21085940 Review.
References
-
- Ayscough, K. R., J. J. Eby, T. Lila, H. Dewar, K. G. Kozminski, and D. G. Drubin. 1999. Sla1p is a functionally modular component of the yeast cortical actin cytoskeleton required for correct localization of both Rho1p-GTPase and Sla2p, a protein with talin homology. Mol. Biol. Cell 10:1061-1075. - PMC - PubMed
-
- Balaban, R. S., S. Nemoto, and T. Finkel. 2005. Mitochondria, oxidants, and aging. Cell 120:483-495. - PubMed
-
- Chevtzoff, C., J. Vallortigara, N. Averet, M. Rigoulet, and A. Devin. 2005. The yeast cAMP protein kinase Tpk3p is involved in the regulation of mitochondrial enzymatic content during growth. Biochim. Biophys. Acta 1706:117-125. - PubMed
-
- Choi, J., H. D. Rees, S. T. Weintraub, A. I. Levey, L. S. Chin, and L. Li. 2005. Oxidative modifications and aggregation of Cu, Zn-superoxide dismutase associated with Alzheimer and Parkinson diseases. J. Biol. Chem. 280:11648-11655. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous