

FBPs are calibrated molecular tools to adjust gene expression
- PMID: 16914741
- PMCID: PMC1592819
- DOI: 10.1128/MCB.00754-06
FBPs are calibrated molecular tools to adjust gene expression
Abstract
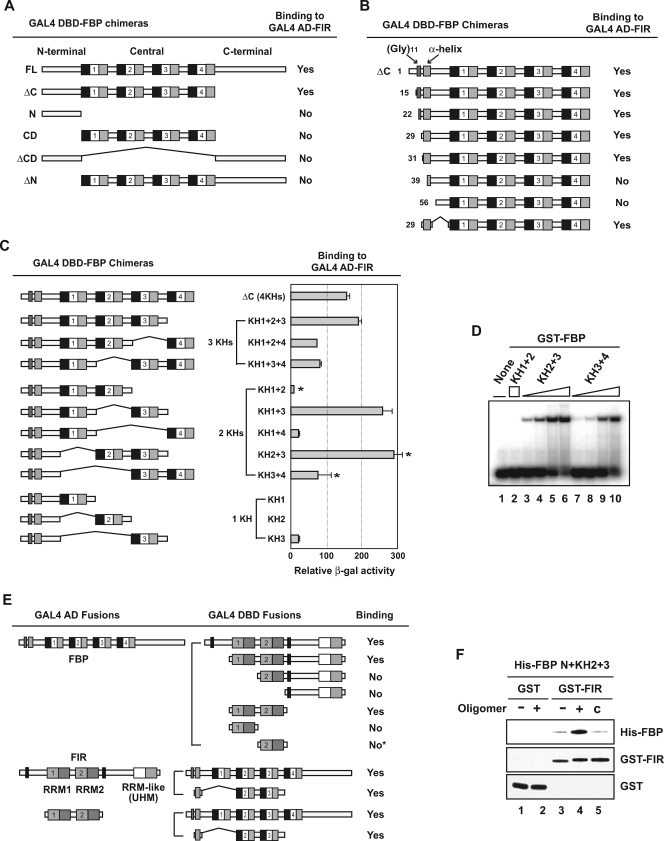
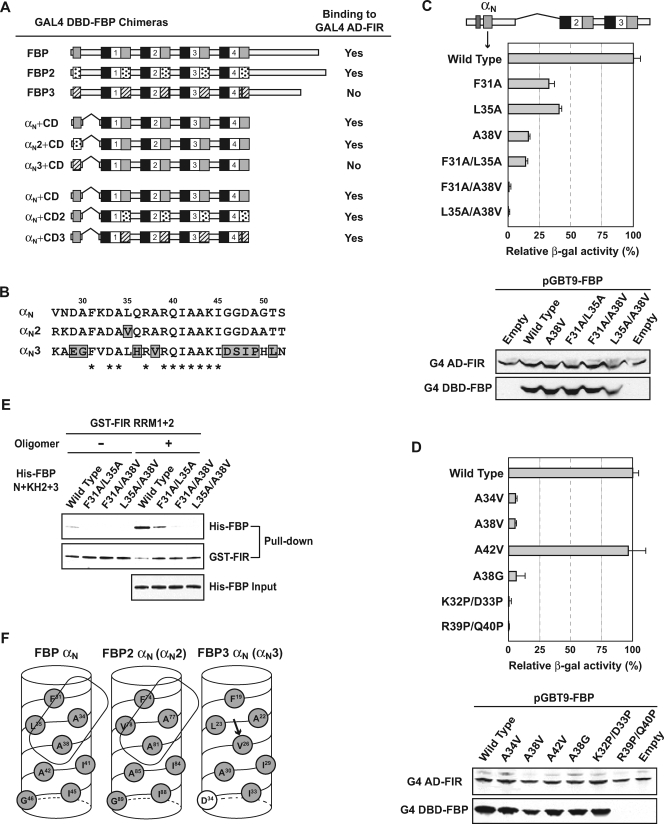
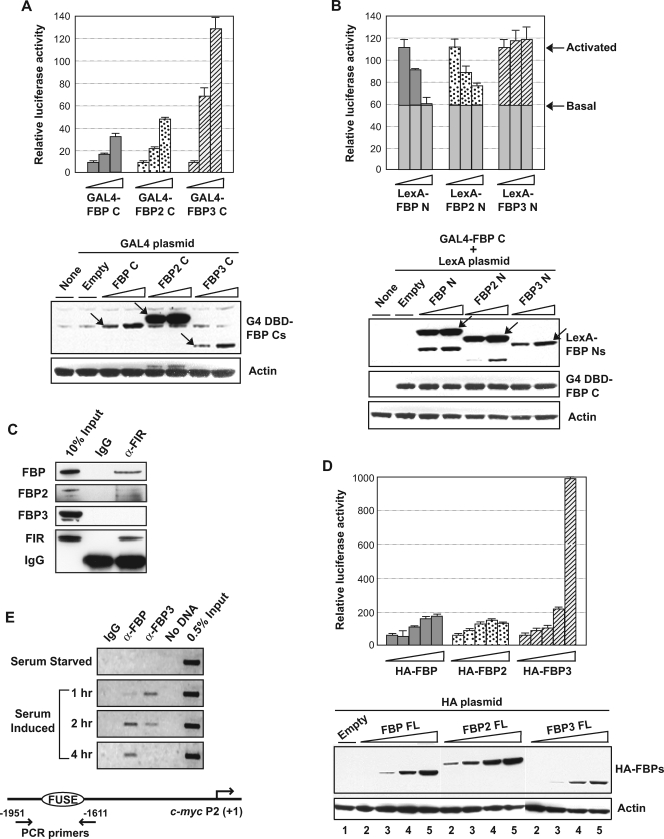
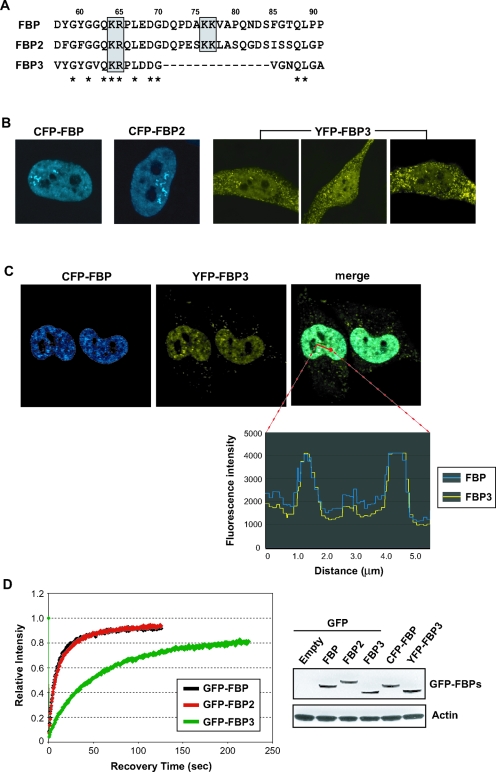
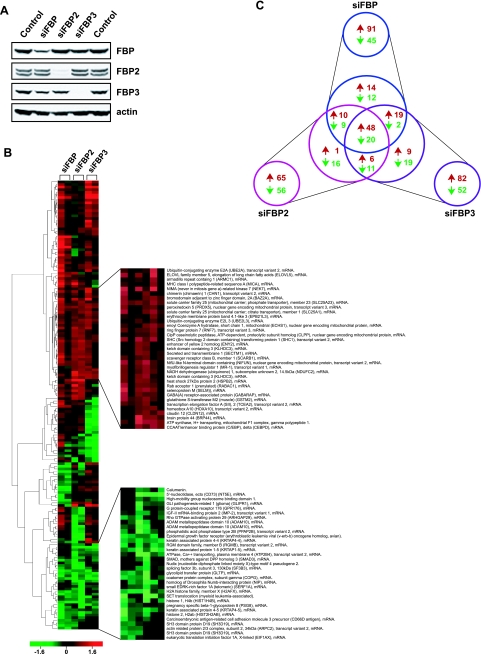
The three far-upstream element (FUSE) binding protein (FBP) family members have been ascribed different functions in gene regulation. They were therefore examined with various biochemical, molecular biological, and cell biological tests to evaluate whether their sequence differences reflect functional customization or neutral changes at unselected residues. Each FBP displayed a characteristic profile of intrinsic transcription activation and repression, binding with protein partners, and subcellular trafficking. Although some differences, such as weakened FBP3 nuclear localization, were predictable from primary sequence differences, the unexpected failure of FBP3 to bind the FBP-interacting repressor (FIR) was traced to seemingly conservative substitutions within a small patch of an N-terminal alpha-helix. The transactivation strength and the FIR-binding strength of the FBPs were in the opposite order. Despite their distinguishing features and differential activities, the FBPs traffic to shared subnuclear sites and regulate many common target genes, including c-myc. Though a variety of functions have been attributed to the FBPs, based upon their panel of shared and unique features, we propose that they constitute a molecular regulatory kit that tunes the expression of shared targets through a common mechanism.
Figures
References
-
- Avigan, M. I., B. Strober, and D. Levens. 1990. A far upstream element stimulates c-myc expression in undifferentiated leukemia cells. J. Biol. Chem. 265:18538-18545. - PubMed
-
- Bazar, L., V. Harris, I. Sunitha, D. Hartmann, and M. Avigan. 1995. A transactivator of c-myc is coordinately regulated with the proto-oncogene during cellular growth. Oncogene 10:2229-2238. - PubMed
-
- Braddock, D. T., J. M. Louis, J. L. Baber, D. Levens, and G. M. Clore. 2002. Structure and dynamics of KH domains from FBP bound to single-stranded DNA. Nature 415:1051-1056. - PubMed
-
- Briata, P., S. V. Forcales, M. Ponassi, G. Corte, C. Y. Chen, M. Karin, P. L. Puri, and R. Gherzi. 2005. p38-dependent phosphorylation of the mRNA decay-promoting factor KSRP controls the stability of select myogenic transcripts. Mol. Cell 20:891-903. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous