

Sodium channel expression in the ventral posterolateral nucleus of the thalamus after peripheral nerve injury
- PMID: 16916452
- PMCID: PMC1563449
- DOI: 10.1186/1744-8069-2-27
Sodium channel expression in the ventral posterolateral nucleus of the thalamus after peripheral nerve injury
Abstract
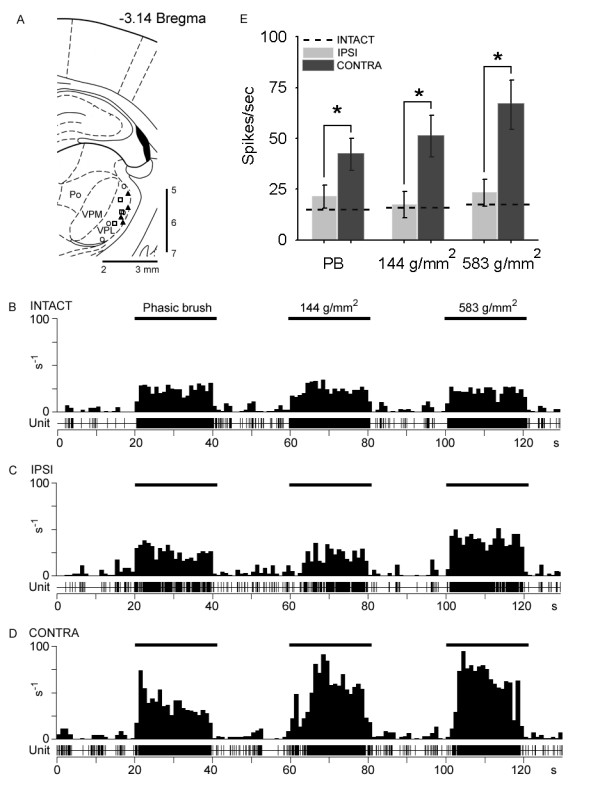
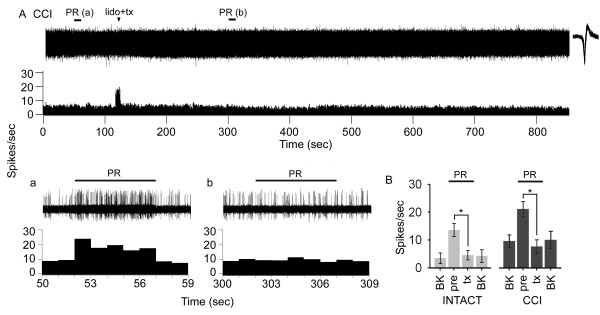
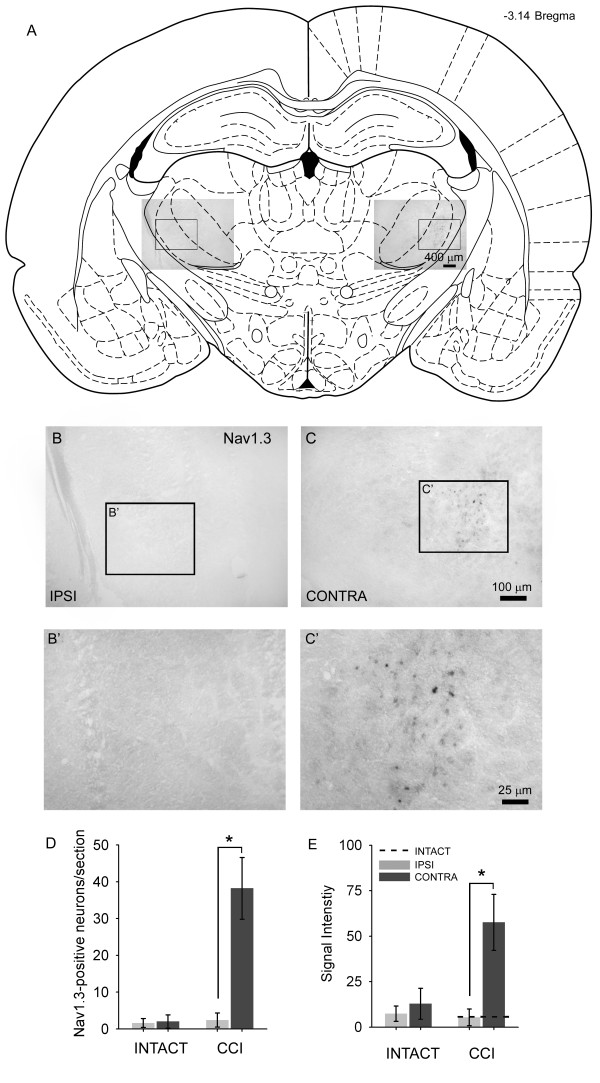
Peripheral nerve injury is known to up-regulate the expression of rapidly-repriming Nav1.3 sodium channel within first-order dorsal root ganglion neurons and second-order dorsal horn nociceptive neurons, but it is not known if pain-processing neurons higher along the neuraxis also undergo changes in sodium channel expression. In this study, we hypothesized that after peripheral nerve injury, third-order neurons in the ventral posterolateral (VPL) nucleus of the thalamus undergo changes in expression of sodium channels. To test this hypothesis, adult male Sprague-Dawley rats underwent chronic constriction injury (CCI) of the sciatic nerve. Ten days after CCI, when allodynia and hyperalgesia were evident, in situ hybridization and immunocytochemical analysis revealed up-regulation of Nav1.3 mRNA, but no changes in expression of Nav1.1, Nav1.2, or Nav1.6 in VPL neurons, and unit recordings demonstrated increased background firing, which persisted after spinal cord transection, and evoked hyperresponsiveness to peripheral stimuli. These results demonstrate that injury to the peripheral nervous system induces alterations in sodium channel expression within higher-order VPL neurons, and suggest that misexpression of the Nav1.3 sodium channel increases the excitability of VPL neurons injury, contributing to neuropathic pain.
Figures

Similar articles
-
Changes in electrophysiological properties and sodium channel Nav1.3 expression in thalamic neurons after spinal cord injury.Brain. 2005 Oct;128(Pt 10):2359-71. doi: 10.1093/brain/awh623. Epub 2005 Aug 18. Brain. 2005. PMID: 16109750
-
Altered sodium channel expression in second-order spinal sensory neurons contributes to pain after peripheral nerve injury.J Neurosci. 2004 May 19;24(20):4832-9. doi: 10.1523/JNEUROSCI.0300-04.2004. J Neurosci. 2004. PMID: 15152043 Free PMC article.
-
Alterations in burst firing of thalamic VPL neurons and reversal by Na(v)1.3 antisense after spinal cord injury.J Neurophysiol. 2006 Jun;95(6):3343-52. doi: 10.1152/jn.01009.2005. Epub 2006 Feb 15. J Neurophysiol. 2006. PMID: 16481457
-
Sodium channel expression and the molecular pathophysiology of pain after SCI.Prog Brain Res. 2007;161:195-203. doi: 10.1016/S0079-6123(06)61013-3. Prog Brain Res. 2007. PMID: 17618978 Review.
-
Distribution of the tetrodotoxin-resistant sodium channel PN3 in rat sensory neurons in normal and neuropathic conditions.J Neurosci. 1998 Mar 15;18(6):2174-87. doi: 10.1523/JNEUROSCI.18-06-02174.1998. J Neurosci. 1998. PMID: 9482802 Free PMC article. Review.
Cited by
-
Neuroplasticity of ascending and descending pathways after somatosensory system injury: reviewing knowledge to identify neuropathic pain therapeutic targets.Spinal Cord. 2016 May;54(5):330-40. doi: 10.1038/sc.2015.225. Epub 2016 Jan 12. Spinal Cord. 2016. PMID: 26754470 Review.
-
The effect of intravenous lidocaine on brain activation during non-noxious and acute noxious stimulation of the forepaw: a functional magnetic resonance imaging study in the rat.Anesth Analg. 2009 Jan;108(1):334-44. doi: 10.1213/ane.0b013e31818e0d34. Anesth Analg. 2009. PMID: 19095870 Free PMC article.
-
Synaptic plasticity in the anterior cingulate cortex in acute and chronic pain.Nat Rev Neurosci. 2016 Aug;17(8):485-96. doi: 10.1038/nrn.2016.68. Epub 2016 Jun 16. Nat Rev Neurosci. 2016. PMID: 27307118 Review.
-
Voltage gated sodium channels as therapeutic targets for chronic pain.J Pain Res. 2019 Sep 9;12:2709-2722. doi: 10.2147/JPR.S207610. eCollection 2019. J Pain Res. 2019. PMID: 31564962 Free PMC article. Review.
-
Tactile responses of hindpaw, forepaw and whisker neurons in the thalamic ventrobasal complex of anesthetized rats.Eur J Neurosci. 2008 Jan;27(2):378-87. doi: 10.1111/j.1460-9568.2008.06025.x. Epub 2008 Jan 11. Eur J Neurosci. 2008. PMID: 18190520 Free PMC article.
References
-
- Sotgiu ML, Lacerenza M, Marchettini P. Effect of systemic lidocaine on dorsal horn neuron hyperactivity following chronic peripheral nerve injury in rats. Somatosens Mot Res. 1992;9:227–233. - PubMed
-
- Laird JM, Bennett GJ. An electrophysiological study of dorsal horn neurons in the spinal cord of rats with an experimental peripheral neuropathy. J Neurophysiol. 1993;69:2072–2085. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
