

Integrin-linked kinase, a novel component of the cardiac mechanical stretch sensor, controls contractility in the zebrafish heart
- PMID: 16921028
- PMCID: PMC1560411
- DOI: 10.1101/gad.1448306
Integrin-linked kinase, a novel component of the cardiac mechanical stretch sensor, controls contractility in the zebrafish heart
Abstract
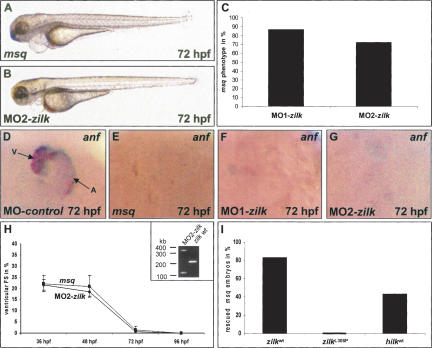
The vertebrate heart possesses autoregulatory mechanisms enabling it first to sense and then to adapt its force of contraction to continually changing demands. The molecular components of the cardiac mechanical stretch sensor are mostly unknown but of immense medical importance, since dysfunction of this sensing machinery is suspected to be responsible for a significant proportion of human heart failure. In the hearts of the ethylnitros-urea (ENU)-induced, recessive embryonic lethal zebrafish heart failure mutant main squeeze (msq), we find stretch-responsive genes such as atrial natriuretic factor (anf) and vascular endothelial growth factor (vegf) severely down-regulated. We demonstrate through positional cloning that heart failure in msq mutants is due to a mutation in the integrin-linked kinase (ilk) gene. ILK specifically localizes to costameres and sarcomeric Z-discs. The msq mutation (L308P) reduces ILK kinase activity and disrupts binding of ILK to the Z-disc adaptor protein beta-parvin (Affixin). Accordingly, in msq mutant embryos, heart failure can be suppressed by expression of ILK, and also of a constitutively active form of Protein Kinase B (PKB), and VEGF. Furthermore, antisense-mediated abrogation of zebrafish beta-parvin phenocopies the msq phenotype. Thus, we provide evidence that the heart uses the Integrin-ILK-beta-parvin network to sense mechanical stretch and respond with increased expression of ANF and VEGF, the latter of which was recently shown to augment cardiac force by increasing the heart's calcium transients.
Figures




Comment in
-
Stretching to meet needs: integrin-linked kinase and the cardiac pump.Genes Dev. 2006 Sep 1;20(17):2327-31. doi: 10.1101/gad.1472506. Genes Dev. 2006. PMID: 16951248 Review. No abstract available.
References
-
- Bock-Marquette I., Saxena A., White M.D., Dimaio J.M., Srivastava D. Thymosin β4 activates integrin-linked kinase and promotes cardiac cell migration, survival and cardiac repair. Nature. 2004;432:466–472. - PubMed
-
- Brancaccio M., Fratta L., Notte A., Hirsch E., Poulet R., Guazzone S., De Acetis M., Vecchione C., Marino G., Altruda F., et al. Melusin, a muscle-specific integrin β1-interacting protein, is required to prevent cardiac failure in response to chronic pressure overload. Nat. Med. 2003;9:68–75. - PubMed
-
- Burggren W.W., Pinder A.W. Ontogeny of cardiovascular and respiratory physiology in lower vertebrates. Annu. Rev. Physiol. 1991;53:107–135. - PubMed
-
- Chan J., Bayliss P.E., Wood J.M., Roberts T.M. Dissection of angiogenic signaling in zebrafish using a chemical genetic approach. Cancer Cell. 2002;1:257–267. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous