

Emi1 stably binds and inhibits the anaphase-promoting complex/cyclosome as a pseudosubstrate inhibitor
- PMID: 16921029
- PMCID: PMC1560415
- DOI: 10.1101/gad.1454006
Emi1 stably binds and inhibits the anaphase-promoting complex/cyclosome as a pseudosubstrate inhibitor
Abstract
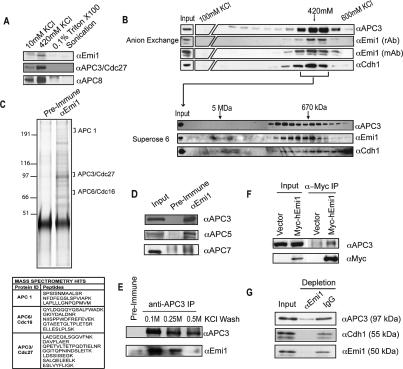
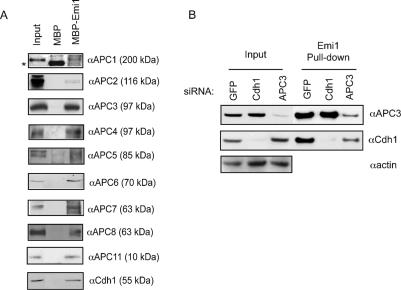
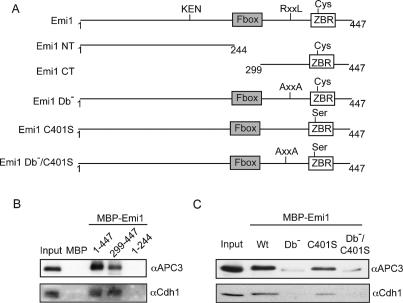
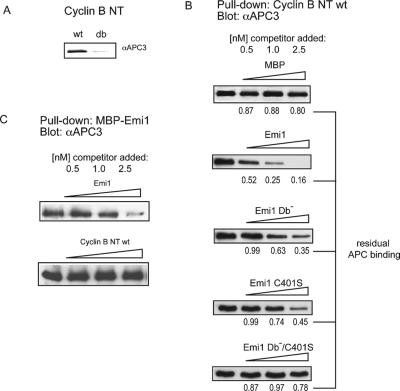
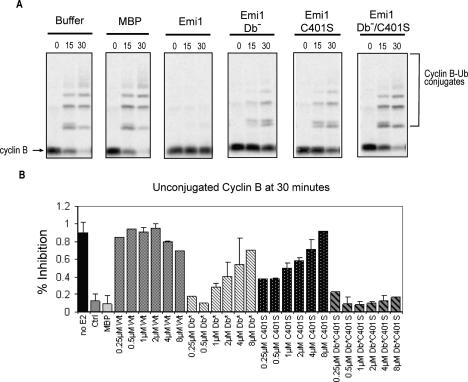
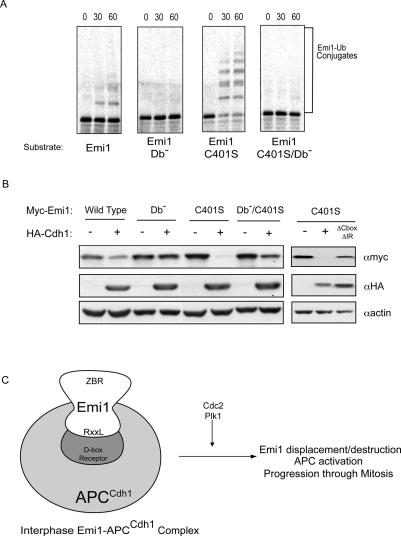
The periodic destruction of mitotic cyclins is triggered by the activation of the anaphase-promoting complex/cyclosome (APC/C) in mitosis. Although the ability of the APC/C to recognize destruction box (D-box) substrates oscillates throughout the cell cycle, the mechanism regulating APC/C binding to D-box substrates remains unclear. Here, we show that the APC/C inhibitor Emi1 tightly binds both the APC/C and its Cdh1 activator, binds to the D-box receptor site on the APC/C(Cdh1), and competes with APC/C substrates for D-box binding. Emi1 itself contains a conserved C-terminal D-box, which provides APC/C-binding affinity, and a conserved zinc-binding region (ZBR), which antagonizes APC/C E3 ligase activity independent of tight APC binding. Mutation of the ZBR converts Emi1 into a D-box-dependent APC/C substrate. The identification of a direct Emi1-APC/C complex further explains how Emi1 functions as a stabilizing factor for cyclin accumulation and the need to destroy Emi1 for APC/C activation in mitosis. The combination of a degron/E3 recognition site and an anti-ligase function in Emi1 suggests a general model for how E3 substrates evolve to become pseudosubstrate inhibitors.
Figures
References
-
- Bashir T., Dorrello N.V., Amador V., Guardavaccaro D., Pagano M. Control of the SCF(Skp2–Cks1) ubiquitin ligase by the APC/C(Cdh1) ubiquitin ligase. Nature. 2004;428:190–193. - PubMed
-
- Burton J.L., Tsakraklides V., Solomon M.J. Assembly of an APC–Cdh1–substrate complex is stimulated by engagement of a destruction box. Mol. Cell. 2005;18:533–542. - PubMed
-
- Carroll C.W., Enquist-Newman M., Morgan D.O. The APC subunit Doc1 promotes recognition of the substrate destruction box. Curr. Biol. 2005;15:11–18. - PubMed
-
- Castro A., Bernis C., Vigneron S., Labbe J.C., Lorca T. The anaphase-promoting complex: A key factor in the regulation of cell cycle. Oncogene. 2005;24:314–325. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous