

Discordance in the effects of Yersinia pestis on the dendritic cell functions manifested by induction of maturation and paralysis of migration
- PMID: 16923789
- PMCID: PMC1695518
- DOI: 10.1128/IAI.00974-06
Discordance in the effects of Yersinia pestis on the dendritic cell functions manifested by induction of maturation and paralysis of migration
Abstract
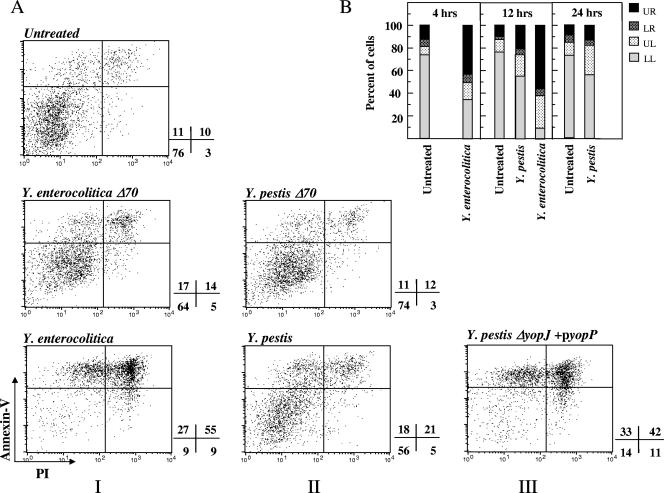
The encounter between invading microorganisms and dendritic cells (DC) triggers a series of events which include uptake and degradation of the microorganism, induction of a maturation process, and enhancement of DC migration to the draining lymph nodes. Various pathogens have developed strategies to counteract these events as a measure to evade the host defense. In the present study we found that interaction of the Yersinia pestis EV76 strain with DC has no effect on cell viability and is characterized by compliance with effective maturation, which is manifested by surface display of major histocompatibility complex class II, of costimulatory markers, and of the chemokine receptor CCR7. This is in contrast to maturation inhibition and cell death induction exerted by the related species Yersinia enterocolitica WA O:8. Y. pestis interactions with DC were found, however, to impair functions related to cytoskeleton rearrangement. DC pulsed with Y. pestis failed to adhere to solid surfaces and to migrate toward the chemokine CCL19 in an in vitro transmembrane assay. Both effects were dependent on the presence of the pCD1 virulence plasmid and on a bacterial growth shift to 37 degrees C prior to infection. Moreover, while instillation of a pCD1-cured Y. pestis strain into mouse airways triggered effective transport of alveolar DC to the mediastinal lymph node, instillation of Y. pestis harboring the plasmid failed to do so. Taken together, these results suggest that virulence plasmid-dependent impairment of DC migration is the major mechanism utilized by Y. pestis to subvert DC function.
Figures
Similar articles
-
Yersinia pestis evades TLR4-dependent induction of IL-12(p40)2 by dendritic cells and subsequent cell migration.J Immunol. 2008 Oct 15;181(8):5560-7. doi: 10.4049/jimmunol.181.8.5560. J Immunol. 2008. PMID: 18832714 Free PMC article.
-
Yersinia enterocolitica induces apoptosis and inhibits surface molecule expression and cytokine production in murine dendritic cells.Infect Immun. 2004 Dec;72(12):7045-54. doi: 10.1128/IAI.72.12.7045-7054.2004. Infect Immun. 2004. PMID: 15557628 Free PMC article.
-
Yersinia pestis subverts the dermal neutrophil response in a mouse model of bubonic plague.mBio. 2013 Aug 27;4(5):e00170-13. doi: 10.1128/mBio.00170-13. mBio. 2013. PMID: 23982068 Free PMC article.
-
[Intestinal yersinia infection: pathogenesis, clinical course, epidemiology and diagnosis].Immun Infekt. 1982 Sep;10(5):180-6. Immun Infekt. 1982. PMID: 6757111 Review. German.
-
Coordinated events during bacteria-induced DC maturation.Immunol Today. 1999 May;20(5):200-3. doi: 10.1016/s0167-5699(98)01427-3. Immunol Today. 1999. PMID: 10322296 Review. No abstract available.
Cited by
-
Yersinia pestis endowed with increased cytotoxicity is avirulent in a bubonic plague model and induces rapid protection against pneumonic plague.PLoS One. 2009 Jun 16;4(6):e5938. doi: 10.1371/journal.pone.0005938. PLoS One. 2009. PMID: 19529770 Free PMC article.
-
Tn5AraOut mutagenesis for the identification of Yersinia pestis genes involved in resistance towards cationic antimicrobial peptides.Microb Pathog. 2011 Sep;51(3):121-32. doi: 10.1016/j.micpath.2011.04.010. Epub 2011 May 7. Microb Pathog. 2011. PMID: 21575704 Free PMC article.
-
Haemophilus ducreyi partially activates human myeloid dendritic cells.Infect Immun. 2007 Dec;75(12):5678-85. doi: 10.1128/IAI.00702-07. Epub 2007 Oct 8. Infect Immun. 2007. PMID: 17923525 Free PMC article.
-
Interaction between Yersinia pestis and the host immune system.Infect Immun. 2008 May;76(5):1804-11. doi: 10.1128/IAI.01517-07. Epub 2008 Feb 4. Infect Immun. 2008. PMID: 18250178 Free PMC article. Review. No abstract available.
-
Gr1+ cells control growth of YopM-negative yersinia pestis during systemic plague.Infect Immun. 2009 Sep;77(9):3791-806. doi: 10.1128/IAI.00284-09. Epub 2009 Jul 6. Infect Immun. 2009. PMID: 19581396 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
