

Exo-oligosaccharides of Rhizobium sp. strain NGR234 are required for symbiosis with various legumes
- PMID: 16923883
- PMCID: PMC1595362
- DOI: 10.1128/JB.00365-06
Exo-oligosaccharides of Rhizobium sp. strain NGR234 are required for symbiosis with various legumes
Erratum in
-
Correction for Staehelin et al., "Exo-Oligosaccharides of Rhizobium sp. Strain NGR234 Are Required for Symbiosis with Various Legumes".J Bacteriol. 2025 Aug 20:e0015725. doi: 10.1128/jb.00157-25. Online ahead of print. J Bacteriol. 2025. PMID: 40833097 No abstract available.
Abstract
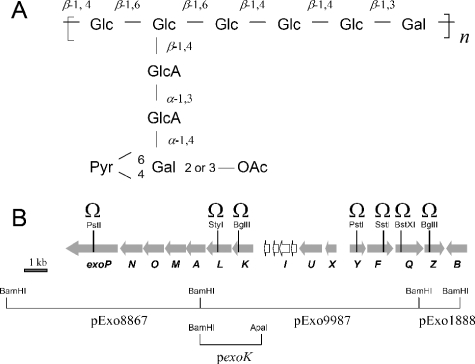
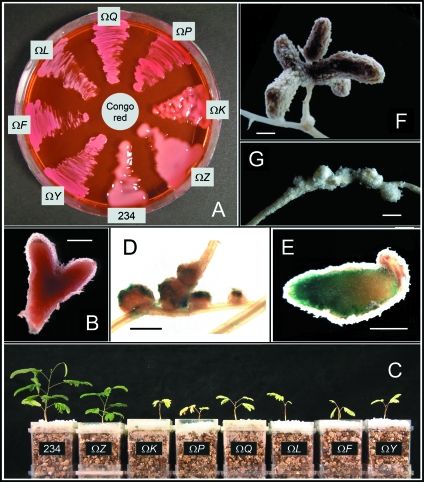
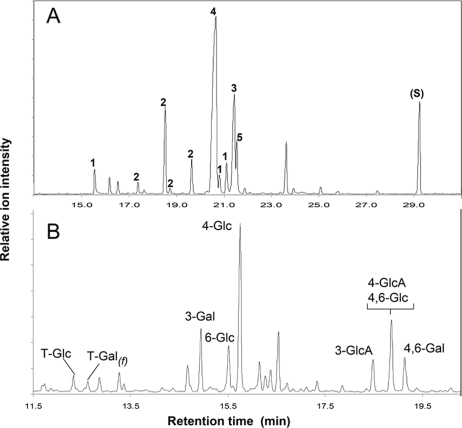
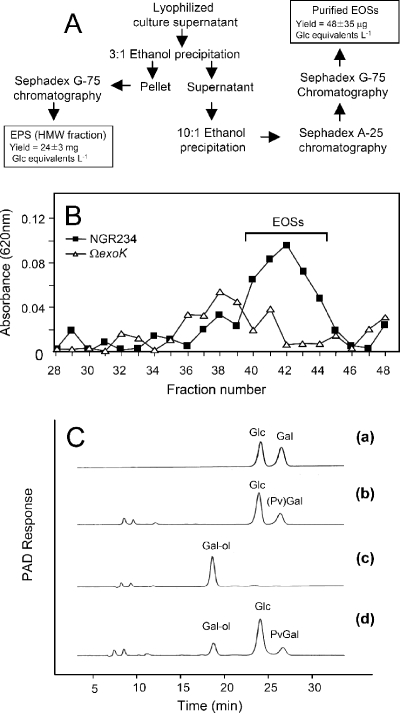
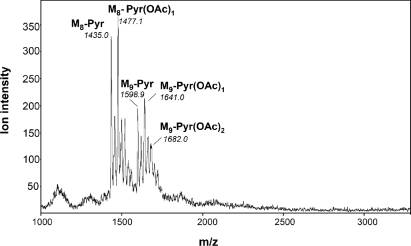
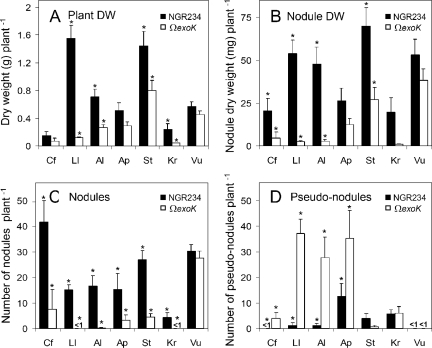
Rhizobia are nitrogen-fixing bacteria that establish endosymbiotic associations with legumes. Nodule formation depends on various bacterial carbohydrates, including lipopolysaccharides, K-antigens, and exopolysaccharides (EPS). An acidic EPS from Rhizobium sp. strain NGR234 consists of glucosyl (Glc), galactosyl (Gal), glucuronosyl (GlcA), and 4,6-pyruvylated galactosyl (PvGal) residues with beta-1,3, beta-1,4, beta-1,6, alpha-1,3, and alpha-1,4 glycoside linkages. Here we examined the role of NGR234 genes in the synthesis of EPS. Deletions within the exoF, exoL, exoP, exoQ, and exoY genes suppressed accumulation of EPS in bacterial supernatants, a finding that was confirmed by chemical analyses. The data suggest that the repeating subunits of EPS are assembled by an ExoQ/ExoP/ExoF-dependent mechanism, which is related to the Wzy polymerization system of group 1 capsular polysaccharides in Escherichia coli. Mutation of exoK (NGROmegaexoK), which encodes a putative glycanase, resulted in the absence of low-molecular-weight forms of EPS. Analysis of the extracellular carbohydrates revealed that NGROmegaexoK is unable to accumulate exo-oligosaccharides (EOSs), which are O-acetylated nonasaccharide subunits of EPS having the formula Gal(Glc)5(GlcA)2PvGal. When used as inoculants, both the exo-deficient mutants and NGROmegaexoK were unable to form nitrogen-fixing nodules on some hosts (e.g., Albizia lebbeck and Leucaena leucocephala), but they were able to form nitrogen-fixing nodules on other hosts (e.g., Vigna unguiculata). EOSs of the parent strain were biologically active at very low levels (yield in culture supernatants, approximately 50 microg per liter). Thus, NGR234 produces symbiotically active EOSs by enzymatic degradation of EPS, using the extracellular endo-beta-1,4-glycanase encoded by exoK (glycoside hydrolase family 16). We propose that the derived EOSs (and not EPS) are bacterial components that play a crucial role in nodule formation in various legumes.
Figures
Similar articles
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
Sexual Harassment and Prevention Training.2024 Mar 29. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Mar 29. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 36508513 Free Books & Documents.
-
Heterologous exopolysaccharide production in Rhizobium sp. strain NGR234 and consequences for nodule development.J Bacteriol. 1991 May;173(10):3066-77. doi: 10.1128/jb.173.10.3066-3077.1991. J Bacteriol. 1991. PMID: 2022612 Free PMC article.
-
Drugs for preventing postoperative nausea and vomiting in adults after general anaesthesia: a network meta-analysis.Cochrane Database Syst Rev. 2020 Oct 19;10(10):CD012859. doi: 10.1002/14651858.CD012859.pub2. Cochrane Database Syst Rev. 2020. PMID: 33075160 Free PMC article.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
Cited by
-
Rhizobial migration toward roots mediated by FadL-ExoFQP modulation of extracellular long-chain AHLs.ISME J. 2023 Mar;17(3):417-431. doi: 10.1038/s41396-023-01357-5. Epub 2023 Jan 10. ISME J. 2023. PMID: 36627434 Free PMC article.
-
Naringenin regulates expression of genes involved in cell wall synthesis in Herbaspirillum seropedicae.Appl Environ Microbiol. 2011 Mar;77(6):2180-3. doi: 10.1128/AEM.02071-10. Epub 2011 Jan 21. Appl Environ Microbiol. 2011. PMID: 21257805 Free PMC article.
-
Symbiotic use of pathogenic strategies: rhizobial protein secretion systems.Nat Rev Microbiol. 2009 Apr;7(4):312-20. doi: 10.1038/nrmicro2091. Epub 2009 Mar 9. Nat Rev Microbiol. 2009. PMID: 19270720 Review.
-
Symbiosis-promoting and deleterious effects of NopT, a novel type 3 effector of Rhizobium sp. strain NGR234.J Bacteriol. 2008 Jul;190(14):5101-10. doi: 10.1128/JB.00306-08. Epub 2008 May 16. J Bacteriol. 2008. PMID: 18487326 Free PMC article.
-
Exopolysaccharide biosynthesis enables mature biofilm formation on abiotic surfaces by Herbaspirillum seropedicae.PLoS One. 2014 Oct 13;9(10):e110392. doi: 10.1371/journal.pone.0110392. eCollection 2014. PLoS One. 2014. PMID: 25310013 Free PMC article.
References
-
- Becker, A., and A. Pühler. 1998. Production of exopolysaccharides, p. 97-118. In H. P. Spaink, Á. Kondorosi, and P. J. J. Hooykaas (ed.), The Rhizobiaceae. Kluwer Academic Publishers, Dordrecht, The Netherlands.
-
- Becker, A., N. Fraysse, and L. Sharypova. 2005. Recent advances in studies on structure and symbiosis-related function of rhizobial K-antigens and lipopolysaccharides. Mol. Plant-Microbe Interact. 9:899-905. - PubMed
-
- Broughton, W. J., and C. K. John. 1979. Rhizobia in tropical legumes. III. Experimentation and supply in Malaysia 1927-1976, p. 113-136. In W. J. Broughton, C. K. John, B. Lim, and C. Rajova (ed.), Soil microbiology and plant nutrition. University of Malaysia, Kuala Lumpur, Malaysia.
-
- Chen, H., M. Batley, J. W. Redmond, and B. G. Rolfe. 1985. Alteration of the effective nodulation properties of a fast-growing broad host-range Rhizobium due to changes in exopolysaccharide synthesis. J. Plant Physiol. 120:331-349.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
