

doi: 10.1128/JB.00668-06.
Genetic evidence for an interaction of the UbiG O-methyltransferase with UbiX in Escherichia coli coenzyme Q biosynthesis
Affiliations
- PMID: 16923914
- PMCID: PMC1595381
- DOI: 10.1128/JB.00668-06
Item in Clipboard
Genetic evidence for an interaction of the UbiG O-methyltransferase with UbiX in Escherichia coli coenzyme Q biosynthesis
J Bacteriol.
2006 Sep.
Abstract
IS16 is a thiol-sensitive, Q-deficient mutant strain of Escherichia coli. Here, we show that IS16 harbors a mutation in the ubiG gene encoding a methyltransferase required for two O-methylation steps of Q biosynthesis. Complementation of IS16 with either ubiG or ubiX(K-12) reverses this phenotype, suggesting that UbiX may interact with UbiG.
Figures

Alignment of E. coli UbiG, S. cerevisiae Coq3, human Coq3, and rat COMT amino acid sequences across methyltransferase motifs I, post-I, II, and III. The box designates the L132Q ubiG mutation in the IS16 mutant strain. This amino acid substitution is adjacent to methyltransferase motif II. Sequence analysis revealed that ubiG genes from THU and IS16 share five nucleotide variances in comparison to the ubiG gene from K-12. These variances include T23A, resulting in V8E, and four silent changes, G90T, C109T, A294G, and C321T. Secondary structural elements (β1, β2, β4, and β5) and important active site residues involved in the binding of ligands are indicated for the rat soluble COMT. a, AdoMet; m, magnesium; s, substrate (27).
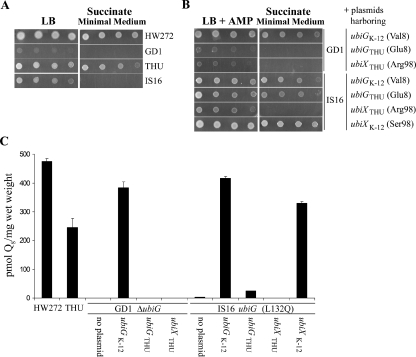
Succinate growth and Q8 levels in E. coli strains. (A) Serial 10-fold dilutions (starting optical density at 600 nm, 0.2) of the indicated strains were plated on LB and succinate minimal medium plates supplemented with thymine, histidine, and uracil and incubated at 37°C for 1 day. (B) Serial 10-fold dilutions (starting optical density at 600 nm, 0.2) of the indicated strains were plated on LB-AMP and succinate minimal medium plates supplemented with thymine, histidine, and uracil and incubated at 37°C for 1 day. E. coli strains harbored plasmids expressing the designated genes as follows: pAHG, UbiGK-12; pUbiGTHU, UbiGTHU; pHZ1, UbiXTHU; and pPZ2, UbiXK-12. (C) Quinones were extracted and separated by reverse-phase HPLC, and Q8 content was quantified. A standard for Q8 was prepared from E. coli lipid extracts and verified by mass spectrophotometric methods. External standard curves were created for Q8 (600, 300, 75, and 30 pmol) and Q10 (300, 150, and 37.5 pmol). The amounts of Q8 were corrected by recovery of the Q10 internal standard.

O methylation of early Q intermediates is defective in IS16 and is restored by expression of E. coli UbiXK-12. Permeabilized E. coli cells were prepared from HW272 (wild-type, parental strain of GD1), GD1 (ubiG disruption mutant), THU (parental strain of IS16), IS16 (UbiG-L132Q), and IS16:pPZ2 (1S16 harboring UbiXK-12 on a plasmid) and incubated with farnesylated analogs of 1 mM 3,4-dihydroxy-5-farnesylbenzoic acid (open bars), 250 μM 5-farnesyl-2-hydroxyphenol (closed bars) for 10 min. S-Adenosyl-[methyl-3H]l -methionine (6.9 μM, 81.5 Ci/mmol; PerkinElmer Life Sciences) was added to the reaction mixture. Following incubation at 37°C for 45 min, lipids were extracted and separated by reverse-phase HPLC (BetaBasic C18 column, 5 μM, 4.6 by 250 mm; Thermo Electron Corporation). Radioactivity present in fractions 6 and 7 (open bars) or fractions 10 to 12 (closed bars) is expressed in pmol of CH3 groups/hr/mg wet weight and eluted with the 3H-labeled product standard. Error bars represent standard deviations obtained from two O-methyltransferase assays of the same sample, and data shown represent two independent experiments.
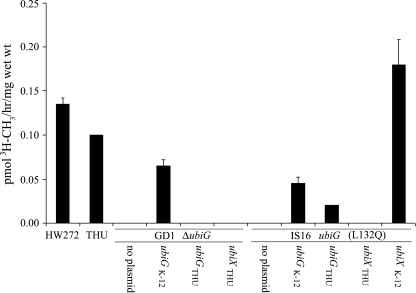
The final O-methylation reaction is defective in the IS16 mutant and is restored by expression of either UbiG or UbiXK-12. In vitro O-methyltransferase assays were carried out as described for Fig. 3, except that 50 μM 2-farnesyl-5-hydroxy-6-methoxy-3-methyl-1,4 benzoquinone was used as a substrate, and 1 mM NADH was included in all incubations to allow formation of the hydroquinone. At the end of the reaction incubation, 25 μl of freshly prepared 1% ammonium cerium (IV) nitrate was added to the reaction mixture prior to lipid extraction (to oxidize hydroquinone products). The elution positions of the methylated products (fractions 11 and 12) were similar to that of the 3H-labeled product standard and are expressed in pmol of CH3 groups/hr/mg wet weight. Error bars represent standard deviations obtained from two O-methyltransferase assays of the same sample, and data shown represent two independent experiments. E. coli strains harbored plasmids expressing the designated genes as follows: pAHG, UbiGK-12; pUbiGTHU, UbiGTHU; pHZ1, UbiXTHU; and pPZ2, UbiXK-12.

Steady-state levels of UbiG and cytochrome o oxidase in E. coli strains. Purified E. coli protein His6-UbiG (24) was used to generate antisera in rabbits (Cocalico Biologicals, Inc.). Steady-state levels of UbiG were analyzed for E. coli HW272 (wild-type, parental strain of GD1), GD1 (ubiG disruption mutant), THU (parental strain of IS16), IS16 (UbiG-L132Q), and IS16:pPZ2 (IS16 complemented with UbiXK-12) cells. Cytochrome o oxidase was used as a loading control.

Genetic evidence for the interaction of UbiX and UbiG. IS16 harbors a mutation in UbiG (L132Q) that catalyzes the O-methylation step in E. coli coenzyme Q biosynthesis. Lack of growth on succinate, Q deficiency, thiol hypersensitivity (29), and inactive O-methyltransferase activity in IS16 are restored with wild-type ubiXK-12, suggesting that UbiX may be interacting with UbiG (L132Q). X, UbiX; G, UbiG.
Similar articles
-
The role of UbiX in Escherichia coli coenzyme Q biosynthesis.Arch Biochem Biophys. 2007 Nov 15;467(2):144-53. doi: 10.1016/j.abb.2007.08.009. Epub 2007 Aug 23. Arch Biochem Biophys. 2007. PMID: 17889824 Free PMC article.
-
Complementation of coq3 mutant yeast by mitochondrial targeting of the Escherichia coli UbiG polypeptide: evidence that UbiG catalyzes both O-methylation steps in ubiquinone biosynthesis.Biochemistry. 1996 Jul 30;35(30):9797-806. doi: 10.1021/bi9602932. Biochemistry. 1996. PMID: 8703953
-
Restoring de novo coenzyme Q biosynthesis in Caenorhabditis elegans coq-3 mutants yields profound rescue compared to exogenous coenzyme Q supplementation.Gene. 2012 Sep 10;506(1):106-16. doi: 10.1016/j.gene.2012.06.023. Epub 2012 Jun 23. Gene. 2012. PMID: 22735617 Free PMC article.
-
Biosynthesis and physiology of coenzyme Q in bacteria.Biochim Biophys Acta. 2014 Jul;1837(7):1004-11. doi: 10.1016/j.bbabio.2014.01.015. Epub 2014 Jan 28. Biochim Biophys Acta. 2014. PMID: 24480387 Review.
-
The UbiX-UbiD system: The biosynthesis and use of prenylated flavin (prFMN).Arch Biochem Biophys. 2017 Oct 15;632:209-221. doi: 10.1016/j.abb.2017.07.014. Epub 2017 Jul 25. Arch Biochem Biophys. 2017. PMID: 28754323 Review.
Cited by
-
Chorismate pyruvate-lyase and 4-hydroxy-3-solanesylbenzoate decarboxylase are required for plastoquinone biosynthesis in the cyanobacterium Synechocystis sp. PCC6803.J Biol Chem. 2014 Jan 31;289(5):2675-86. doi: 10.1074/jbc.M113.511709. Epub 2013 Dec 11. J Biol Chem. 2014. PMID: 24337576 Free PMC article.
-
Biosynthesis of Menaquinone (Vitamin K2) and Ubiquinone (Coenzyme Q).EcoSal Plus. 2009 Aug;3(2):10.1128/ecosalplus.3.6.3.3. doi: 10.1128/ecosalplus.3.6.3.3. EcoSal Plus. 2009. PMID: 26443765 Free PMC article.
-
The role of UbiX in Escherichia coli coenzyme Q biosynthesis.Arch Biochem Biophys. 2007 Nov 15;467(2):144-53. doi: 10.1016/j.abb.2007.08.009. Epub 2007 Aug 23. Arch Biochem Biophys. 2007. PMID: 17889824 Free PMC article.
-
Crystallization and preliminary crystallographic studies of UbiG, an O-methyltransferase from Escherichia coli.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2011 Jun 1;67(Pt 6):727-9. doi: 10.1107/S1744309111014278. Epub 2011 May 26. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2011. PMID: 21636923 Free PMC article.
References
-
- Baba, S. W., G. I. Belogrudov, J. C. Lee, P. T. Lee, J. Strahan, J. N. Shepherd, and C. F. Clarke. 2004. Yeast Coq5 C-methyltransferase is required for stability of other polypeptides involved in coenzyme Q biosynthesis. J. Biol. Chem. 279:10052-10059. - PubMed
-
- Bader, M., W. Muse, D. P. Ballou, C. Gassner, and J. C. Bardwell. 1999. Oxidative protein folding is driven by the electron transport system. Cell 98:217-227. - PubMed
-
- Belogrudov, G. I., P. T. Lee, T. Jonassen, A. Y. Hsu, P. Gin, and C. F. Clarke. 2001. Yeast COQ4 encodes a mitochondrial protein required for coenzyme Q. synthesis. Arch. Biochem. Biophys. 392:48-58. - PubMed
-
- Brandt, U., and B. Trumpower. 1994. The protonmotive Q cycle in mitochondria and bacteria. Crit. Rev. Biochem. Mol. Biol. 29:165-197. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
