

The Yng1p plant homeodomain finger is a methyl-histone binding module that recognizes lysine 4-methylated histone H3
- PMID: 16923967
- PMCID: PMC1636756
- DOI: 10.1128/MCB.00573-06
The Yng1p plant homeodomain finger is a methyl-histone binding module that recognizes lysine 4-methylated histone H3
Abstract
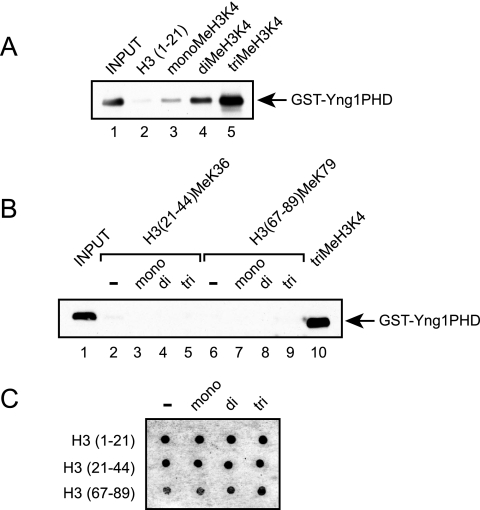
The ING (inhibitor of growth) protein family includes a group of homologous nuclear proteins that share a highly conserved plant homeodomain (PHD) finger domain at their carboxyl termini. Members of this family are found in multiprotein complexes that posttranslationally modify histones, suggesting that these proteins serve a general role in permitting various enzymatic activities to interact with nucleosomes. There are three members of the ING family in Saccharomyces cerevisiae: Yng1p, Yng2p, and Pho23p. Yng1p is a component of the NuA3 histone acetyltransferase complex and is required for the interaction of NuA3 with chromatin. To gain insight into the function of the ING proteins, we made use of a genetic strategy to identify genes required for the binding of Yng1p to histones. Using the toxicity of YNG1 overexpression as a tool, we showed that Yng1p interacts with the amino-terminal tail of histone H3 and that this interaction can be disrupted by loss of lysine 4 methylation within this tail. Additionally, we mapped the region of Yng1p required for overexpression of toxicity to the PHD finger, showed that this region capable of binding lysine 4-methylated histone H3 in vitro, and demonstrated that mutations of the PHD finger that abolish binding in vitro are no longer toxic in vivo. These results identify a novel function for the Yng1p PHD finger in promoting stabilization of the NuA3 complex at chromatin through recognition of histone H3 lysine 4 methylation.
Figures





References
-
- Aasland, R., T. J. Gibson, and A. F. Stewart. 1995. The PHD finger: implications for chromatin-mediated transcriptional regulation. Trends Biochem. Sci. 20:56-59. - PubMed
-
- Ausubel, F. M. 1987. Current protocols in molecular biology. Wiley-Interscience, New York, N.Y.
-
- Carrozza, M. J., B. Li, L. Florens, T. Suganuma, S. K. Swanson, K. K. Lee, W. J. Shia, S. Anderson, J. Yates, M. P. Washburn, and J. L. Workman. 2005. Histone H3 methylation by Set2 directs deacetylation of coding regions by Rpd3S to suppress spurious intragenic transcription. Cell 123:581-592. - PubMed
-
- Choy, J. S., B. T. Tobe, J. H. Huh, and S. J. Kron. 2001. Yng2p-dependent NuA4 histone H4 acetylation activity is required for mitotic and meiotic progression. J. Biol. Chem. 276:43653-43662. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases