

Redox regulation of human OGG1 activity in response to cellular oxidative stress
- PMID: 16923968
- PMCID: PMC1636869
- DOI: 10.1128/MCB.00624-06
Redox regulation of human OGG1 activity in response to cellular oxidative stress
Abstract
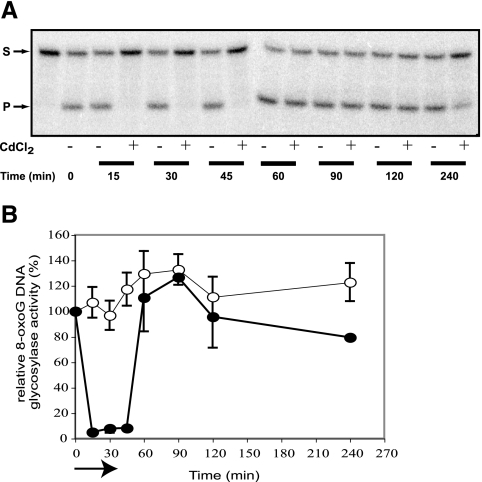
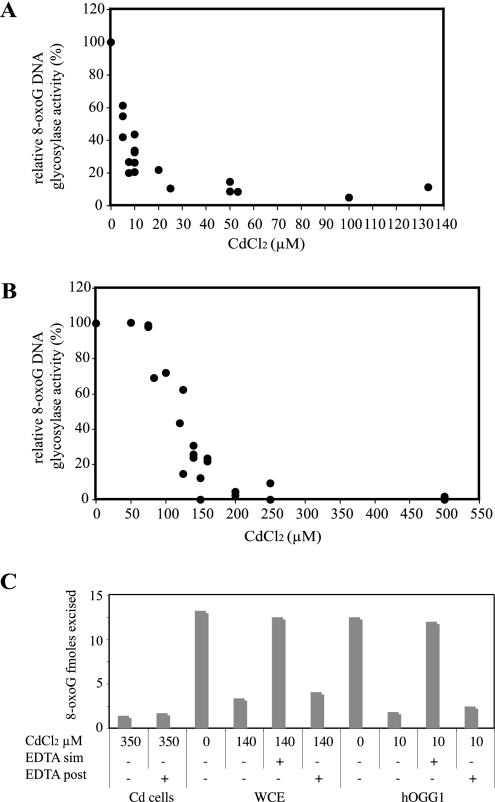
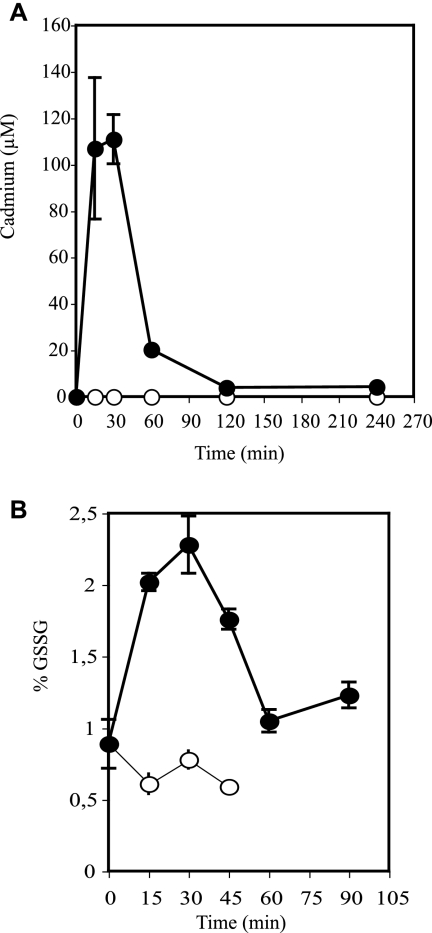
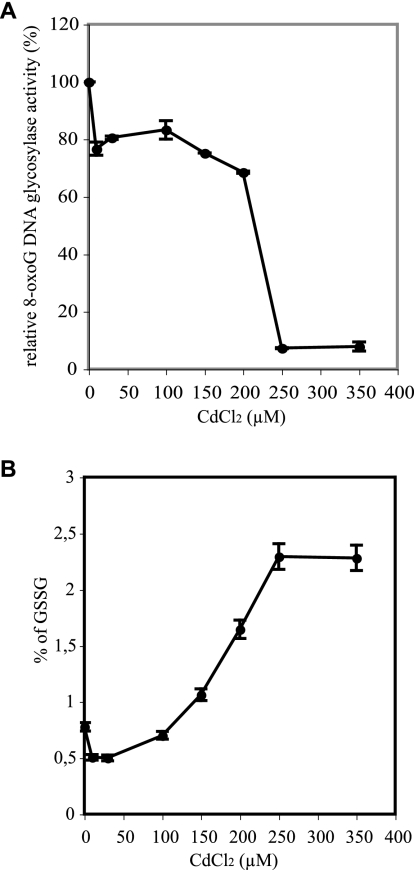
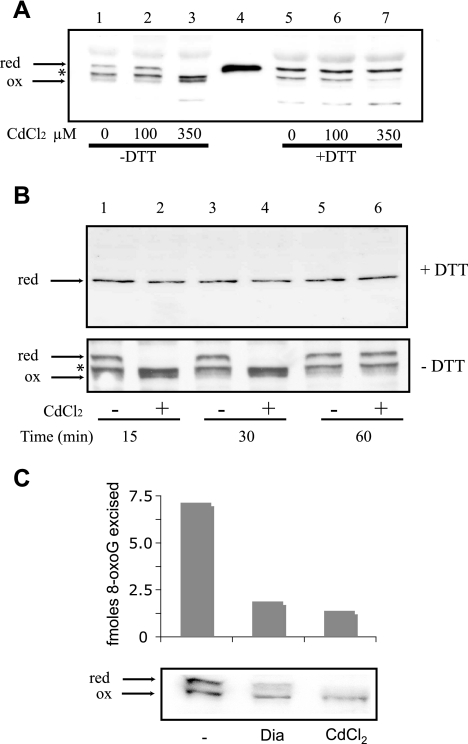
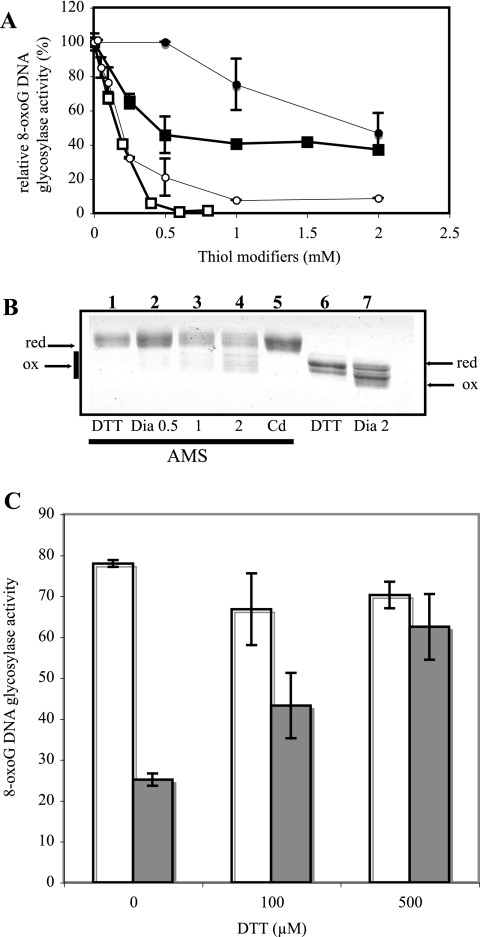
8-Oxoguanine (8-oxoG), a common and mutagenic form of oxidized guanine in DNA, is eliminated mainly through base excision repair. In human cells its repair is initiated by human OGG1 (hOGG1), an 8-oxoG DNA glycosylase. We investigated the effects of an acute cadmium exposure of human lymphoblastoid cells on the activity of hOGG1. We show that coinciding with alteration of the redox cellular status, the 8-oxoG DNA glycosylase activity of hOGG1 was nearly completely inhibited. However, the hOGG1 activity returned to normal levels once the redox cellular status was normalized. In vitro, the activity of purified hOGG1 was abolished by cadmium and could not be recovered by EDTA. In cells, however, the reversible inactivation of OGG1 activity by cadmium was strictly associated with reversible oxidation of the protein. Moreover, the 8-oxoG DNA glycosylase activity of purified OGG1 and that from crude extracts were modulated by cysteine-modifying agents. Oxidation of OGG1 by the thiol oxidant diamide led to inhibition of the activity and a protein migration pattern similar to that seen in cadmium-treated cells. These results suggest that cadmium inhibits hOGG1 activity mainly by indirect oxidation of critical cysteine residues and that excretion of the metal from the cells leads to normalization of the redox cell status and restoration of an active hOGG1. The results presented here unveil a novel redox-dependent mechanism for the regulation of OGG1 activity.
Figures
References
-
- Anderson, M. E. 1985. Tissue glutathione, p. 317-323. In R. A. Greenwald (ed.), Handbook of methods for oxygen radical research. CRC Press, Inc., Boca Raton, Fla.
-
- Audebert, M., J. B. Charbonnier, S. Boiteux, and J. P. Radicella. 2002. Mitochondrial targeting of human 8-oxoguanine DNA glycosylase hOGG1 is impaired by a somatic mutation found in kidney cancer. DNA Repair (Amsterdam) 1:497-505. - PubMed
-
- Biswas, S., A. S. Chida, and I. Rahman. 2006. Redox modifications of protein-thiols: emerging roles in cell signaling. Biochem. Pharmacol. 71:551-564. - PubMed
-
- Boiteux, S., and J. P. Radicella. 2000. The human OGG1 gene: structure, functions, and its implication in the process of carcinogenesis. Arch. Biochem. Biophys. 377:1-8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials