

The DNA polymerase activity of Pol epsilon holoenzyme is required for rapid and efficient chromosomal DNA replication in Xenopus egg extracts
- PMID: 16925818
- PMCID: PMC1560149
- DOI: 10.1186/1471-2091-7-21
The DNA polymerase activity of Pol epsilon holoenzyme is required for rapid and efficient chromosomal DNA replication in Xenopus egg extracts
Abstract
Background: DNA polymerase epsilon (Pol epsilon) is involved in DNA replication, repair, and cell-cycle checkpoint control in eukaryotic cells. Although the roles of replicative Pol alpha and Pol delta in chromosomal DNA replication are relatively well understood and well documented, the precise role of Pol epsilon in chromosomal DNA replication is not well understood.
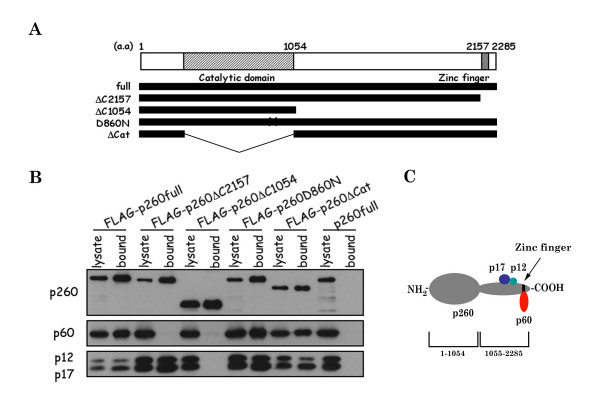
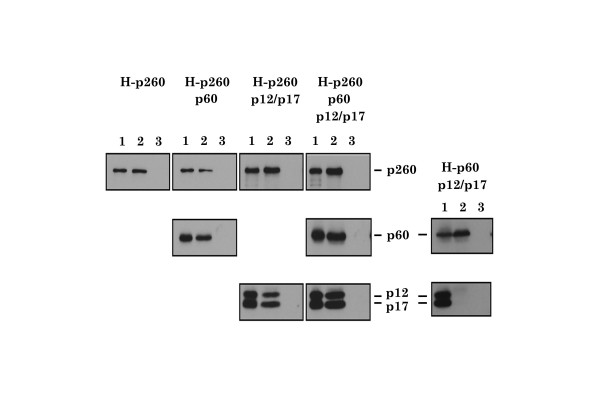
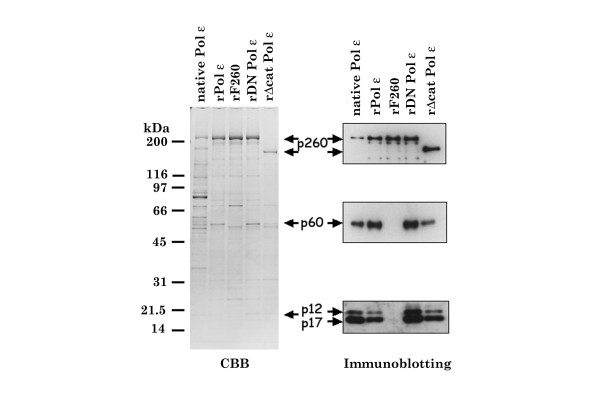
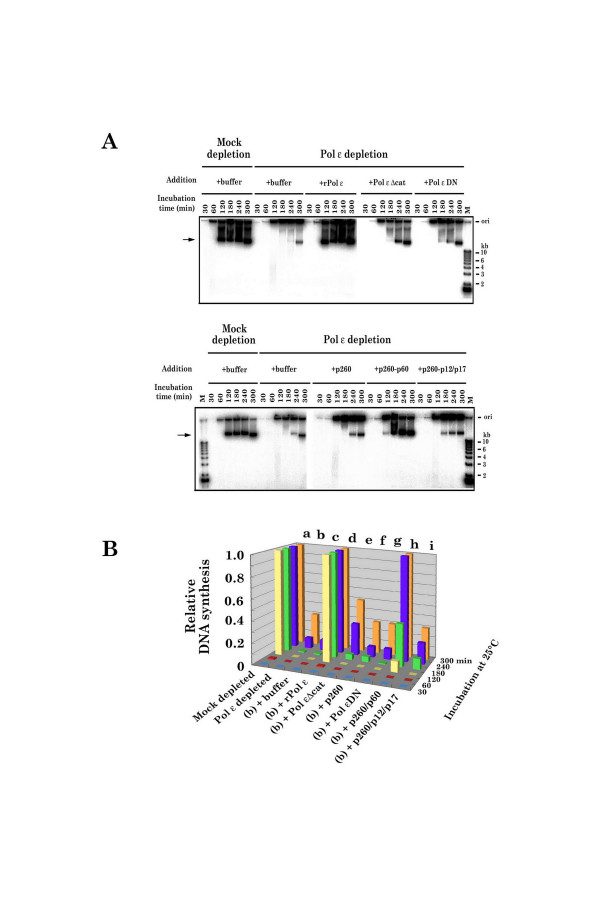
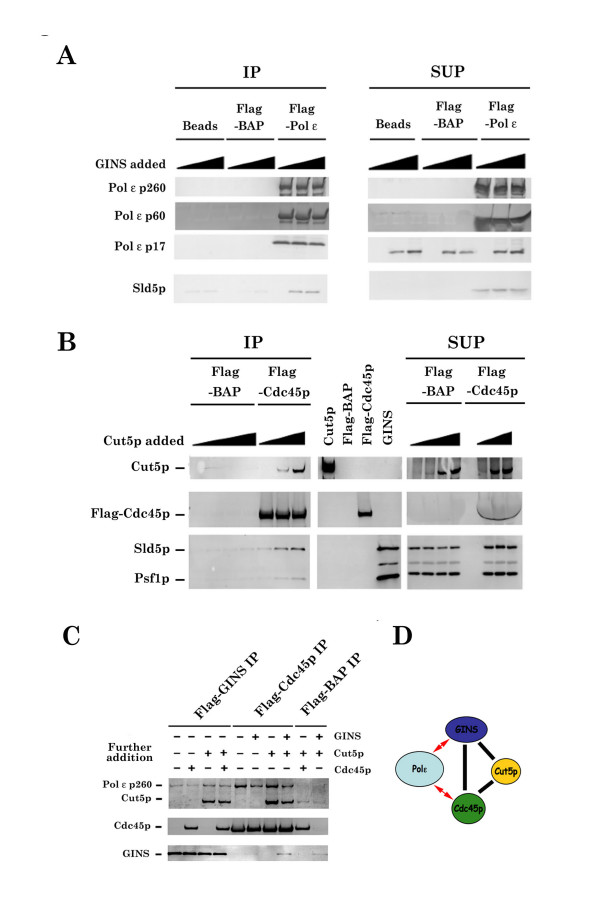
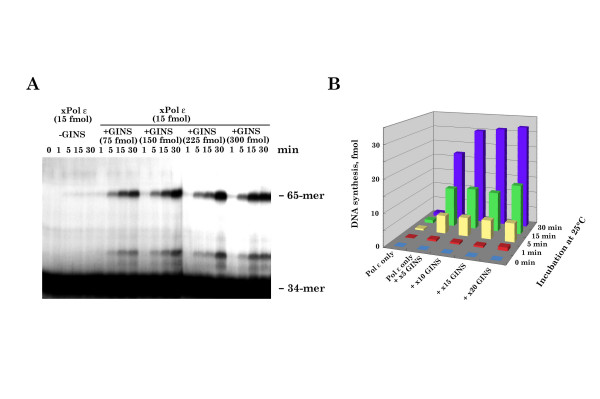
Results: This study uses a Xenopus egg extract DNA replication system to further elucidate the replicative role(s) played by Pol epsilon. Previous studies show that the initiation timing and elongation of chromosomal DNA replication are markedly impaired in Pol epsilon-depleted Xenopus egg extracts, with reduced accumulation of replicative intermediates and products. This study shows that normal replication is restored by addition of Pol epsilon holoenzyme to Pol epsilon-depleted extracts, but not by addition of polymerase-deficient forms of Pol epsilon, including polymerase point or deletion mutants or incomplete enzyme complexes. Evidence is also provided that Pol epsilon holoenzyme interacts directly with GINS, Cdc45p and Cut5p, each of which plays an important role in initiation of chromosomal DNA replication in eukaryotic cells.
Conclusion: These results indicate that the DNA polymerase activity of Pol epsilon holoenzyme plays an essential role in normal chromosomal DNA replication in Xenopus egg extracts. These are the first biochemical data to show the DNA polymerase activity of Pol epsilon holoenzyme is essential for chromosomal DNA replication in higher eukaryotes, unlike in yeasts.
Figures
Similar articles
-
DNA polymerase epsilon is required for coordinated and efficient chromosomal DNA replication in Xenopus egg extracts.Proc Natl Acad Sci U S A. 2001 Apr 24;98(9):4978-83. doi: 10.1073/pnas.081088798. Epub 2001 Apr 10. Proc Natl Acad Sci U S A. 2001. PMID: 11296256 Free PMC article.
-
DNA polymerase epsilon encoded by cdc20+ is required for chromosomal DNA replication in the fission yeast Schizosaccharomyces pombe.Genes Cells. 1998 Feb;3(2):99-110. doi: 10.1046/j.1365-2443.1998.00169.x. Genes Cells. 1998. PMID: 9605404
-
Evidence for interplay among yeast replicative DNA polymerases alpha, delta and epsilon from studies of exonuclease and polymerase active site mutations.BMC Biol. 2004 May 26;2:11. doi: 10.1186/1741-7007-2-11. BMC Biol. 2004. PMID: 15163346 Free PMC article.
-
DNA polymerase ε.Subcell Biochem. 2012;62:237-57. doi: 10.1007/978-94-007-4572-8_13. Subcell Biochem. 2012. PMID: 22918589 Review.
-
Chromosomal DNA replication in a soluble cell-free system derived from Xenopus eggs.Methods Mol Biol. 2006;322:121-37. doi: 10.1007/978-1-59745-000-3_9. Methods Mol Biol. 2006. PMID: 16739720 Review.
Cited by
-
Identification of genes differentially expressed by calorie restriction in the rotifer (Brachionus plicatilis).J Comp Physiol B. 2010 Jan;180(1):105-16. doi: 10.1007/s00360-009-0389-6. Epub 2009 Jul 19. J Comp Physiol B. 2010. PMID: 19618192
-
Crystal structure of the human GINS complex.Genes Dev. 2007 Jun 1;21(11):1316-21. doi: 10.1101/gad.1548107. Genes Dev. 2007. PMID: 17545466 Free PMC article.
-
SV40 DNA replication: from the A gene to a nanomachine.Virology. 2009 Feb 20;384(2):352-9. doi: 10.1016/j.virol.2008.11.038. Epub 2008 Dec 20. Virology. 2009. PMID: 19101707 Free PMC article. Review.
-
Polymerase ε1 mutation in a human syndrome with facial dysmorphism, immunodeficiency, livedo, and short stature ("FILS syndrome").J Exp Med. 2012 Dec 17;209(13):2323-30. doi: 10.1084/jem.20121303. Epub 2012 Dec 10. J Exp Med. 2012. PMID: 23230001 Free PMC article.
-
Reduced expression of GINS complex members induces hallmarks of pre-malignancy in primary untransformed human cells.Cell Cycle. 2009 May 15;8(10):1577-88. doi: 10.4161/cc.8.10.8535. Epub 2009 May 23. Cell Cycle. 2009. PMID: 19377277 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
