

Constitutive expression of outer surface protein C diminishes the ability of Borrelia burgdorferi to evade specific humoral immunity
- PMID: 16926410
- PMCID: PMC1594837
- DOI: 10.1128/IAI.00713-06
Constitutive expression of outer surface protein C diminishes the ability of Borrelia burgdorferi to evade specific humoral immunity
Abstract
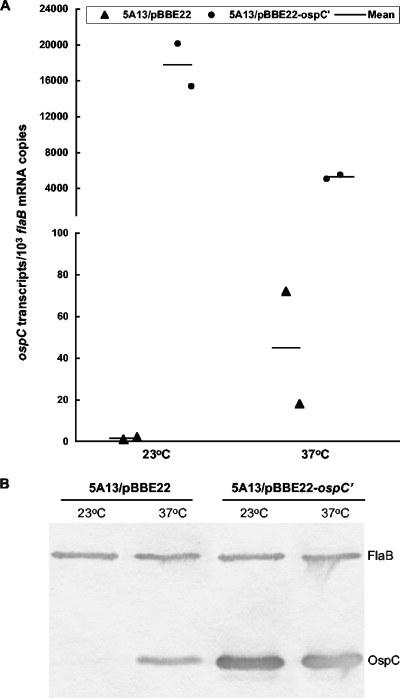
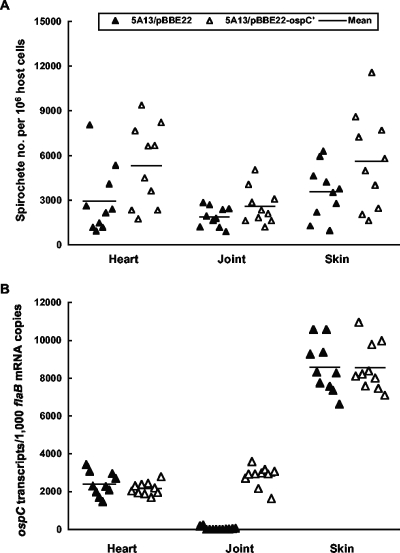
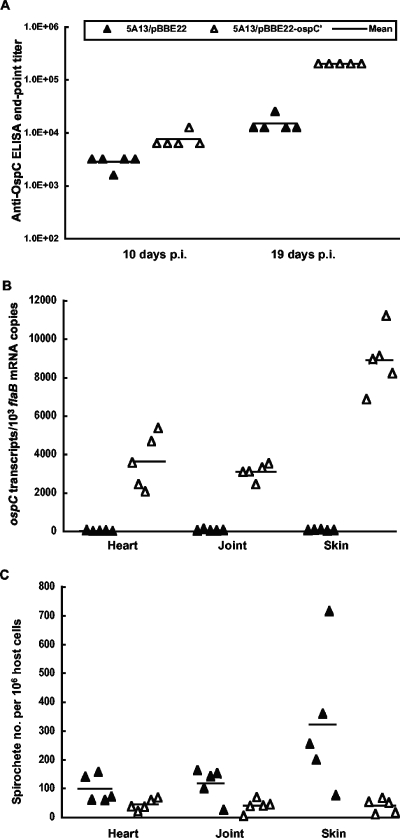
The Lyme disease spirochete Borrelia burgdorferi reduces the expression of outer surface protein C (OspC) in response to the development of an anti-OspC humoral response, leading to the hypothesis that the ability to repress OspC expression is critical for the pathogen to proceed to chronic infection. B. burgdorferi was genetically modified to constitutively express OspC by introducing an extra ospC copy fused with the borrelial flagellar gene (flaB) promoter. Such a genetic modification did not reduce infectivity or pathogenicity in severe combined immunodeficiency mice but resulted in clearance of infection by passively transferred OspC antibody. Spirochetes with constitutive ospC expression were unable to establish chronic infections in immunocompetent mice unless they had undergone very destructive mutations in the introduced ospC copy. Two escape mutants were identified; one had all 7 bp deleted between the putative ribosome-binding site and the start codon, ATG, causing a failure in translational initiation, and the other mutant had an insertion of 2 bp between nucleotides 315 and 316, resulting in a nonsense mutation at codon 108. Thus, the ability of B. burgdorferi to repress ospC expression during mammalian infection allows the pathogen to avoid clearance and to preserve the integrity of the important gene for subsequent utilization during its enzootic life cycle.
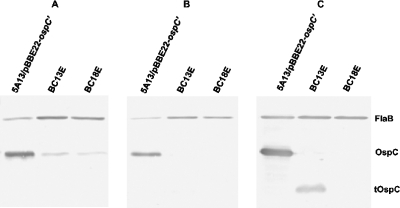
Figures
Similar articles
-
Verification and dissection of the ospC operator by using flaB promoter as a reporter in Borrelia burgdorferi.Microb Pathog. 2008 Jul;45(1):70-8. doi: 10.1016/j.micpath.2008.03.002. Epub 2008 Mar 25. Microb Pathog. 2008. PMID: 18479884 Free PMC article.
-
Identification of an ospC operator critical for immune evasion of Borrelia burgdorferi.Mol Microbiol. 2007 Apr;64(1):220-31. doi: 10.1111/j.1365-2958.2007.05636.x. Mol Microbiol. 2007. PMID: 17376084
-
Outer surface protein OspC is an antiphagocytic factor that protects Borrelia burgdorferi from phagocytosis by macrophages.Infect Immun. 2015 Dec;83(12):4848-60. doi: 10.1128/IAI.01215-15. Epub 2015 Oct 5. Infect Immun. 2015. PMID: 26438793 Free PMC article.
-
Immune evasion of Borrelia burgdorferi: insufficient killing of the pathogens by complement and antibody.Int J Med Microbiol. 2002 Jun;291 Suppl 33:141-6. doi: 10.1016/s1438-4221(02)80027-3. Int J Med Microbiol. 2002. PMID: 12141738 Review.
-
Molecular diversity of the ospC gene in Borrelia. Impact on phylogeny, epidemiology and pathology.Wien Klin Wochenschr. 2002 Jul 31;114(13-14):562-7. Wien Klin Wochenschr. 2002. PMID: 12422602 Review.
Cited by
-
Borrelia burgdorferi spatiotemporal regulation of transcriptional regulator bosR and decorin binding protein during murine infection.Sci Rep. 2020 Jul 27;10(1):12534. doi: 10.1038/s41598-020-69212-7. Sci Rep. 2020. PMID: 32719448 Free PMC article.
-
In Vivo Imaging Demonstrates That Borrelia burgdorferi ospC Is Uniquely Expressed Temporally and Spatially throughout Experimental Infection.PLoS One. 2016 Sep 9;11(9):e0162501. doi: 10.1371/journal.pone.0162501. eCollection 2016. PLoS One. 2016. PMID: 27611840 Free PMC article.
-
Outer surface protein C is a dissemination-facilitating factor of Borrelia burgdorferi during mammalian infection.PLoS One. 2010 Dec 31;5(12):e15830. doi: 10.1371/journal.pone.0015830. PLoS One. 2010. PMID: 21209822 Free PMC article.
-
Essential role of the response regulator Rrp2 in the infectious cycle of Borrelia burgdorferi.Infect Immun. 2008 Sep;76(9):3844-53. doi: 10.1128/IAI.00467-08. Epub 2008 Jun 23. Infect Immun. 2008. PMID: 18573895 Free PMC article.
-
Complement Evasion Contributes to Lyme Borreliae-Host Associations.Trends Parasitol. 2020 Jul;36(7):634-645. doi: 10.1016/j.pt.2020.04.011. Epub 2020 May 23. Trends Parasitol. 2020. PMID: 32456964 Free PMC article. Review.
References
-
- Alexopoulou, L., V. Thomas, M. Schnare, Y. Lobet, J. Anguita, R. T. Schoen, R. Medzhitov, E. Fikrig, and R. A. Flavell. 2002. Hyporesponsiveness to vaccination with Borrelia burgdorferi OspA in humans and in TLR1- and TLR2-deficient mice. Nat. Med. 8:878-884. - PubMed
-
- Alverson, J., S. F. Bundle, C. D. Sohaskey, M. C. Lybecker, and D. S. Samuels. 2003. Transcriptional regulation of the ospAB and ospC promoters from Borrelia burgdorferi. Mol. Microbiol. 48:1665-1677. - PubMed
-
- Burgdorfer, W., and J. E. Keirans. 1983. Ticks and Lyme disease in the United States. Ann. Intern. Med. 99:121. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
