

Mimicking phosphorylation of alphaB-crystallin affects its chaperone activity
- PMID: 16928191
- PMCID: PMC1698675
- DOI: 10.1042/BJ20060981
Mimicking phosphorylation of alphaB-crystallin affects its chaperone activity
Abstract
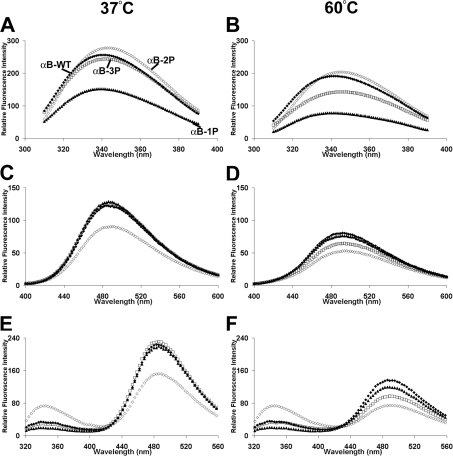
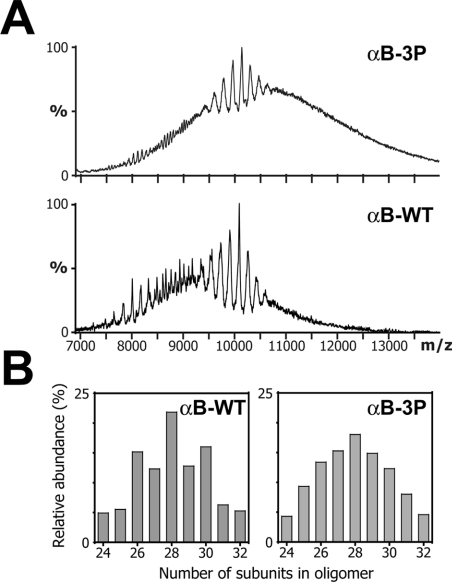
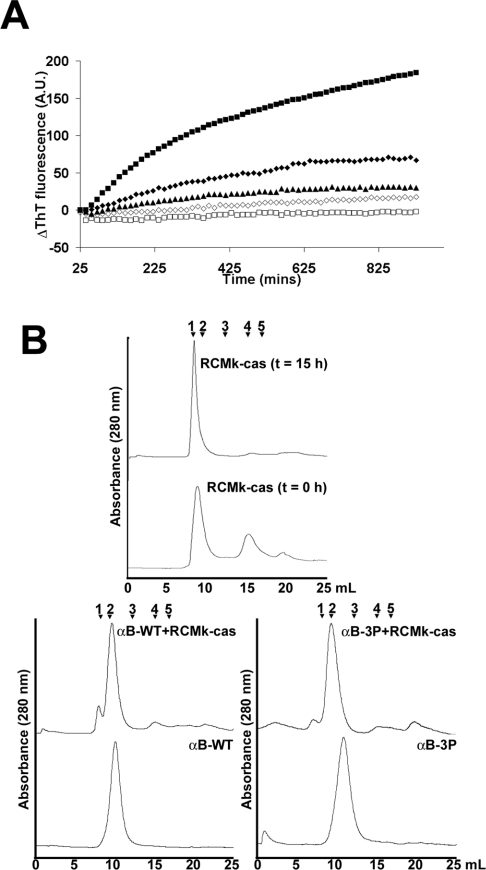
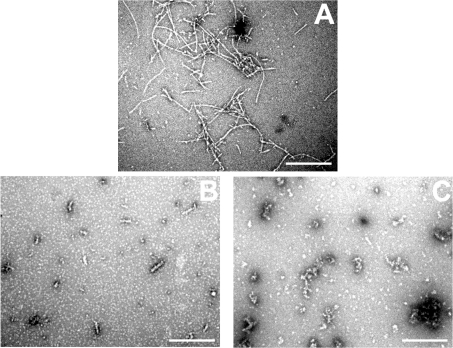
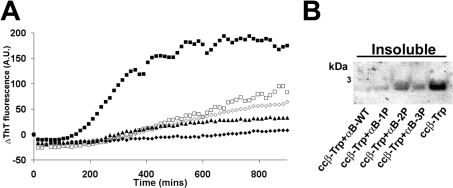
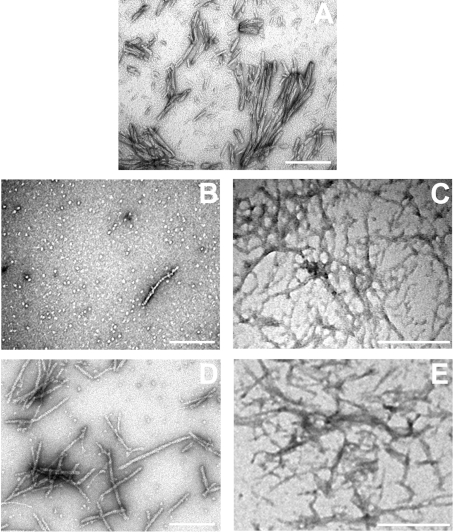
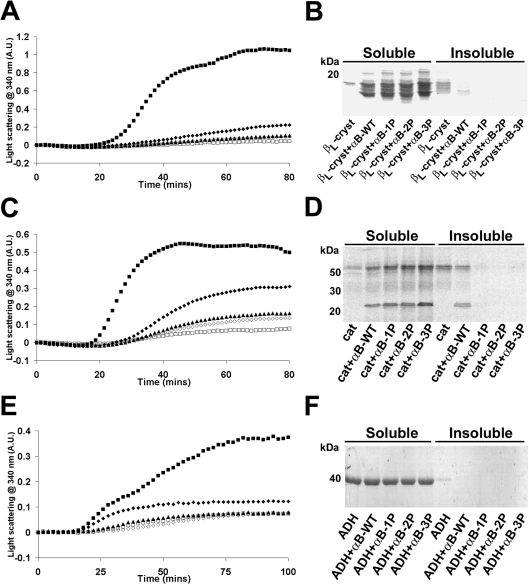
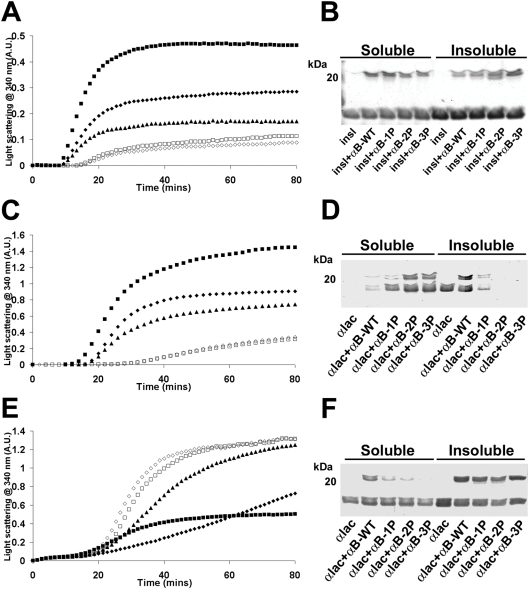
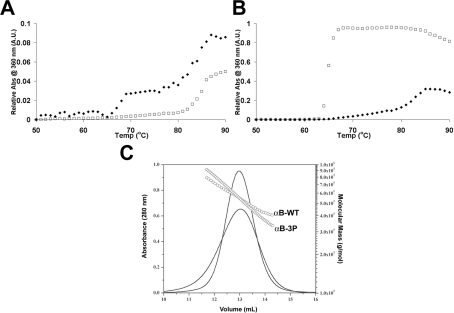
AlphaB-crystallin is a member of the sHsp (small heat-shock protein) family that prevents misfolded target proteins from aggregating and precipitating. Phosphorylation at three serine residues (Ser19, Ser45 and Ser59) is a major post-translational modification that occurs to alphaB-crystallin. In the present study, we produced recombinant proteins designed to mimic phosphorylation of alphaB-crystallin by incorporating a negative charge at these sites. We employed these mimics to undertake a mechanistic and structural investigation of the effect of phosphorylation on the chaperone activity of alphaB-crystallin to protect against two types of protein misfolding, i.e. amorphous aggregation and amyloid fibril assembly. We show that mimicking phosphorylation of alphaB-crystallin results in more efficient chaperone activity against both heat-induced and reduction-induced amorphous aggregation of target proteins. Mimick-ing phosphorylation increased the chaperone activity of alphaB-crystallin against one amyloid-forming target protein (kappa-casein), but decreased it against another (ccbeta-Trp peptide). We observed that both target protein identity and solution (buffer) conditions are critical factors in determining the relative chaperone ability of wild-type and phosphorylated alphaB-crystallins. The present study provides evidence for the regulation of the chaperone activity of alphaB-crystallin by phosphorylation and indicates that this may play an important role in alleviating the pathogenic effects associated with protein conformational diseases.
Figures
Similar articles
-
The small heat-shock protein αB-crystallin uses different mechanisms of chaperone action to prevent the amorphous versus fibrillar aggregation of α-lactalbumin.Biochem J. 2012 Dec 15;448(3):343-52. doi: 10.1042/BJ20121187. Biochem J. 2012. PMID: 23005341
-
The effect of small molecules in modulating the chaperone activity of alphaB-crystallin against ordered and disordered protein aggregation.FEBS J. 2008 Mar;275(5):935-47. doi: 10.1111/j.1742-4658.2008.06257.x. Epub 2008 Jan 23. FEBS J. 2008. PMID: 18218039
-
Enhanced molecular chaperone activity of the small heat-shock protein alphaB-cystallin following covalent immobilization onto a solid-phase support.Biopolymers. 2011 Jun;95(6):376-89. doi: 10.1002/bip.21584. Biopolymers. 2011. PMID: 21225714
-
Proteostasis and the Regulation of Intra- and Extracellular Protein Aggregation by ATP-Independent Molecular Chaperones: Lens α-Crystallins and Milk Caseins.Acc Chem Res. 2018 Mar 20;51(3):745-752. doi: 10.1021/acs.accounts.7b00250. Epub 2018 Feb 14. Acc Chem Res. 2018. PMID: 29442498 Review.
-
A Structural View of αB-crystallin Assembly and Amyloid Aggregation.Protein Pept Lett. 2017;24(4):315-321. doi: 10.2174/0929866524666170206122616. Protein Pept Lett. 2017. PMID: 28176658 Review.
Cited by
-
Multiple molecular architectures of the eye lens chaperone αB-crystallin elucidated by a triple hybrid approach.Proc Natl Acad Sci U S A. 2011 Dec 20;108(51):20491-6. doi: 10.1073/pnas.1111014108. Epub 2011 Dec 5. Proc Natl Acad Sci U S A. 2011. PMID: 22143763 Free PMC article.
-
Role of exosomes in malignant glioma: microRNAs and proteins in pathogenesis and diagnosis.Cell Commun Signal. 2020 Aug 3;18(1):120. doi: 10.1186/s12964-020-00623-9. Cell Commun Signal. 2020. PMID: 32746854 Free PMC article. Review.
-
Pseudophosphorylated αB-crystallin is a nuclear chaperone imported into the nucleus with help of the SMN complex.PLoS One. 2013 Sep 4;8(9):e73489. doi: 10.1371/journal.pone.0073489. eCollection 2013. PLoS One. 2013. PMID: 24023879 Free PMC article.
-
Small heat-shock proteins: important players in regulating cellular proteostasis.Cell Mol Life Sci. 2015 Feb;72(3):429-451. doi: 10.1007/s00018-014-1754-5. Epub 2014 Oct 29. Cell Mol Life Sci. 2015. PMID: 25352169 Free PMC article. Review.
-
αB-crystallin: Portrait of a malignant chaperone as a cancer therapeutic target.Pharmacol Ther. 2016 Apr;160:1-10. doi: 10.1016/j.pharmthera.2016.01.012. Epub 2016 Jan 25. Pharmacol Ther. 2016. PMID: 26820756 Free PMC article. Review.
References
-
- Carver J. A., Rekas A., Thorn D. C., Wilson M. R. Small heat-shock proteins and clusterin: intra- and extracellular molecular chaperones with a common mechanism of action and function? IUBMB Life. 2003;55:661–668. - PubMed
-
- Jakob U., Gaestel M., Engel K., Buchner J. Small heat shock proteins are molecular chaperones. J. Biol. Chem. 1993;268:1517–1520. - PubMed
-
- Clark J. I., Muchowski P. J. Small heat-shock proteins and their potential role in human disease. Curr. Opin. Struct. Biol. 2000;10:52–59. - PubMed
-
- Macario A. J., Conway de Macario E. Sick chaperones, cellular stress, and disease. N. Engl. J. Med. 2005;353:1489–1501. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources