

Axon guidance by diffusible chemoattractants: a gradient of netrin protein in the developing spinal cord
- PMID: 16928876
- PMCID: PMC6674364
- DOI: 10.1523/JNEUROSCI.5191-05.2006
Axon guidance by diffusible chemoattractants: a gradient of netrin protein in the developing spinal cord
Abstract
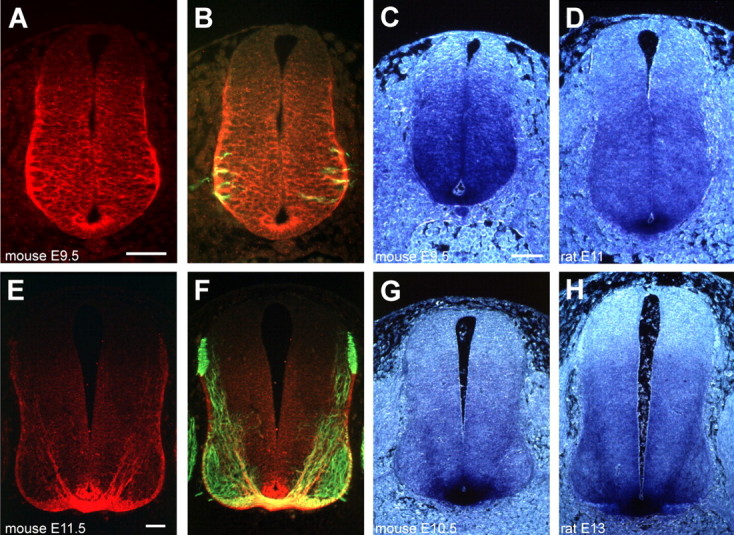
Gradients of diffusible long-range attractant and repellent proteins have been proposed to guide growing axons during nervous system development, but such gradients have never been visualized directly. In the embryonic spinal cord, commissural axons pioneer a circumferential trajectory to the floor plate at the ventral midline directed by secreted proteins of the netrin family. In the embryonic chick spinal cord netrin-1 mRNA is expressed by floor plate cells and netrin-2 mRNA by neural epithelial cells. Antibodies to the two netrins reveal a gradient of netrin protein directly in the path of commissural axons. The netrin-1 gradient itself extends many cell diameters dorsal to the floor plate, the site of netrin-1 expression. A similar distribution of netrin-1 protein has been detected in embryonic rat and mouse spinal cord. The detection of a gradient of netrin-1 protein supports the operation of long-range chemotropic mechanisms in the developing nervous system.
Figures
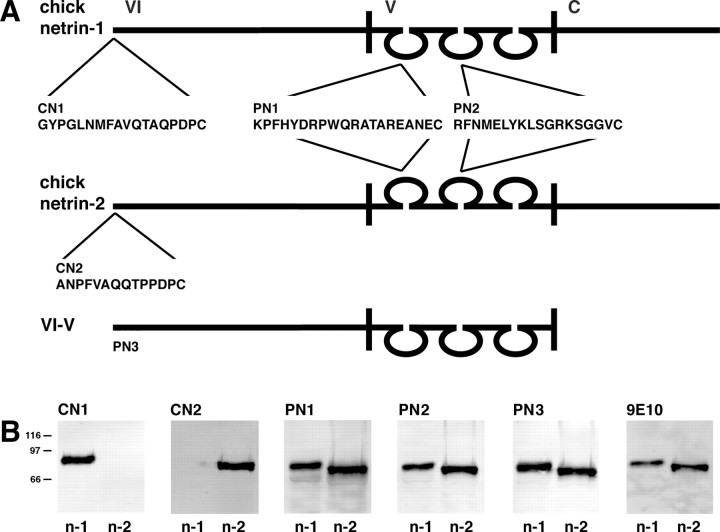
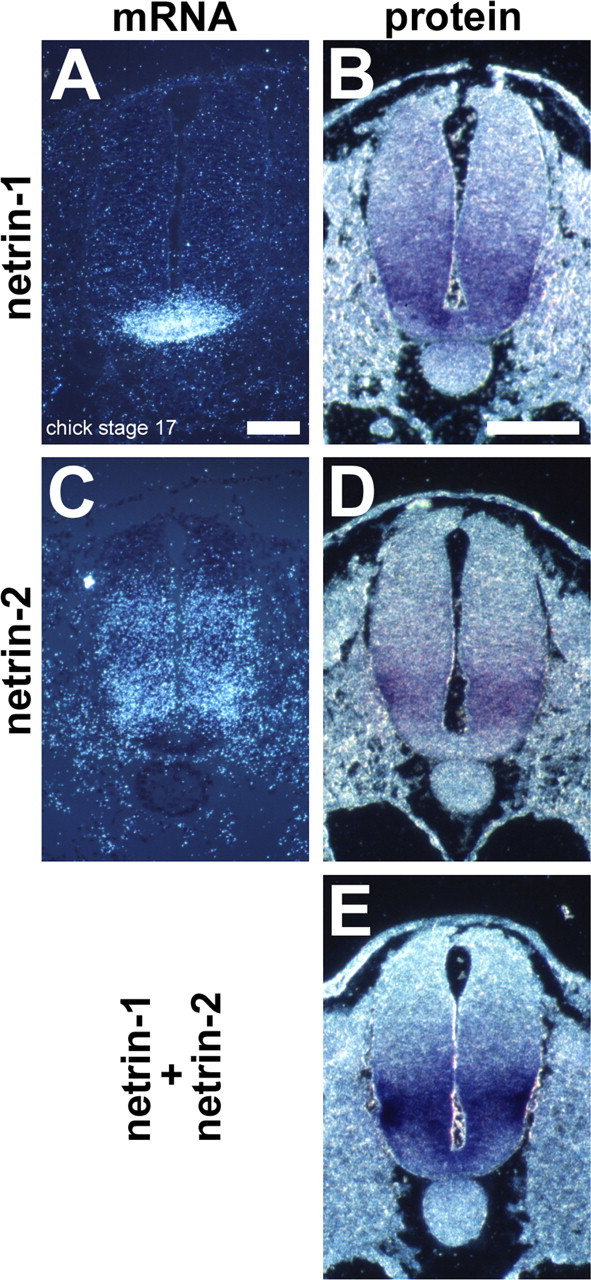
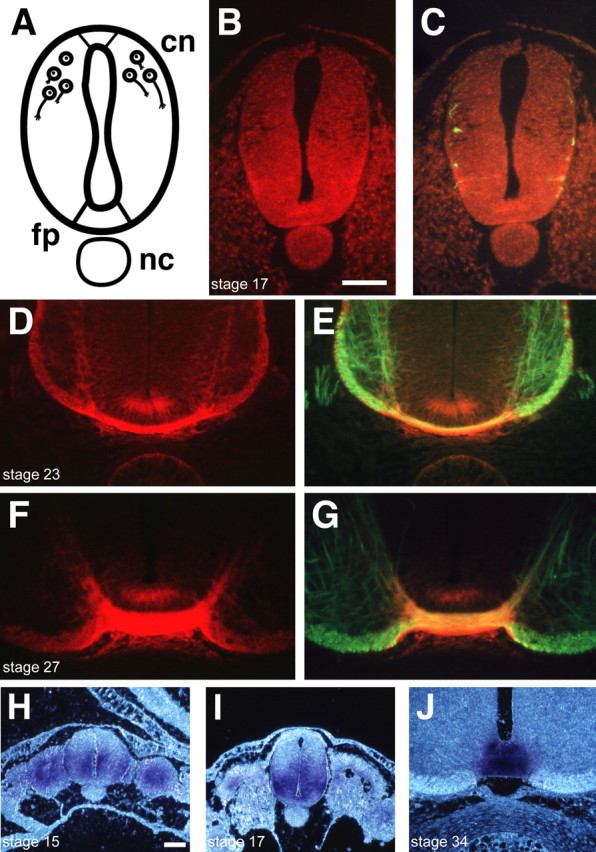
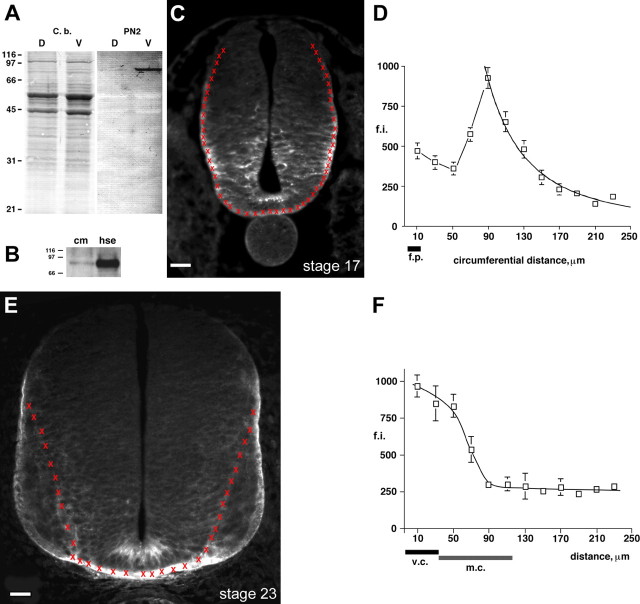
References
-
- Altman J, Bayer SA. The development of the rat spinal cord. Adv Anat Embryol Cell Biol. 1984;85:1–164. - PubMed
-
- Baier H, Bonhoeffer F. Axon guidance by gradients of a target-derived component. Science. 1992;255:472–475. - PubMed
-
- Bourikas D, Pekarik V, Baeriswyl T, Grunditz A, Sadhu R, Nardo M, Stoeckli ET. Sonic hedgehog guides commissural axons along the longitudinal axis of the spinal cord. Nat Neurosci. 2005;8:297–304. - PubMed
-
- Braisted JE, McLaughlin T, Wang HU, Friedman GC, Anderson DJ, O’Leary DD. Graded and lamina-specific distributions of ligands of EphB receptor tyrosine kinases in the developing retinotectal system. Dev Biol. 1997;191:14–28. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases