

Presenilin-dependent intramembrane cleavage of ephrin-B1
- PMID: 16930449
- PMCID: PMC1513130
- DOI: 10.1186/1750-1326-1-2
Presenilin-dependent intramembrane cleavage of ephrin-B1
Abstract
Background: Presenilin-dependent gamma-secretase cleavage of several transmembrane proteins, including amyloid-beta precursor protein and Notch, mediates the intramembrane proteolysis to liberate their intracellular domains that are involved in cellular signaling. Considering gamma-secretase inhibitors as therapeutics for Alzheimer's disease, understanding the physiologically and biologically important substrate for gamma-secretase activity in brains is emerging issue. To elucidate the molecular mechanism and physiological role of gamma-secretase, we screened candidate molecules for gamma-secretase substrates.
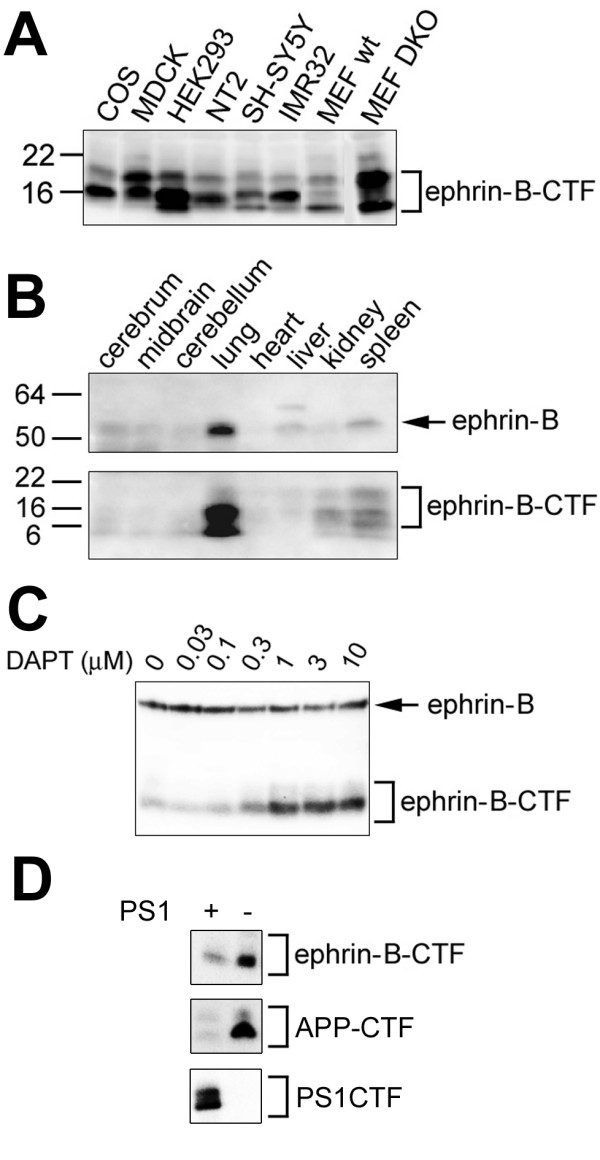
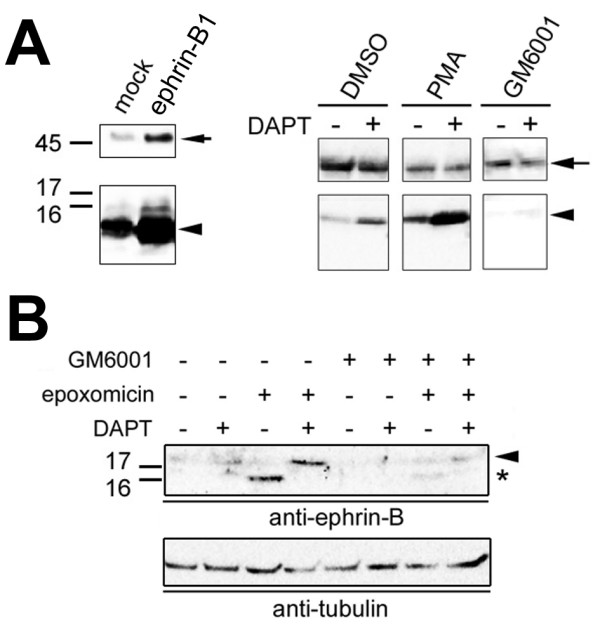
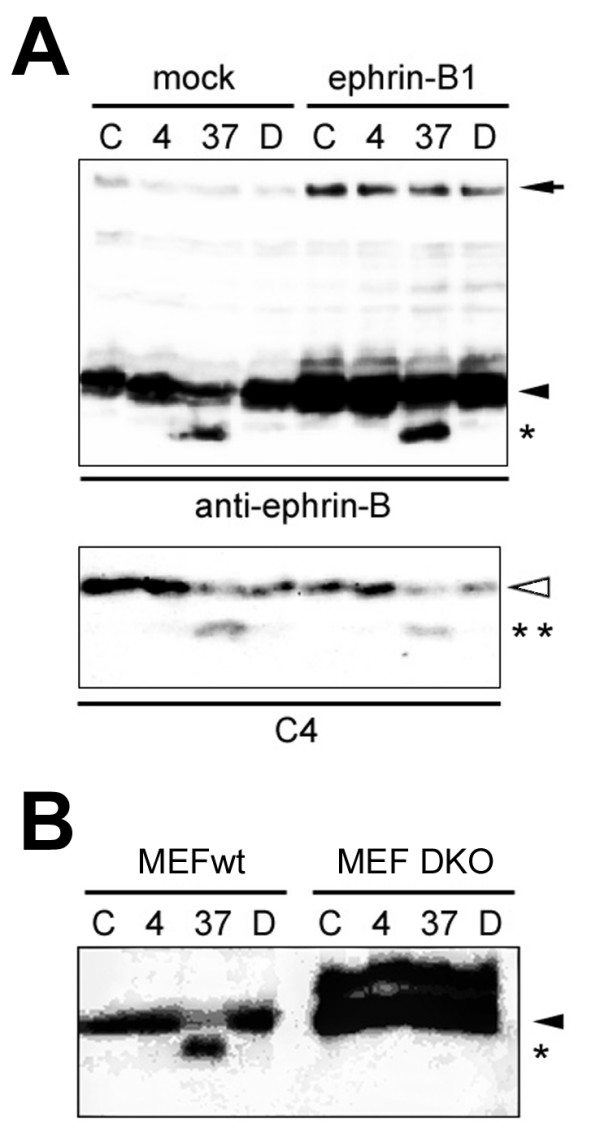
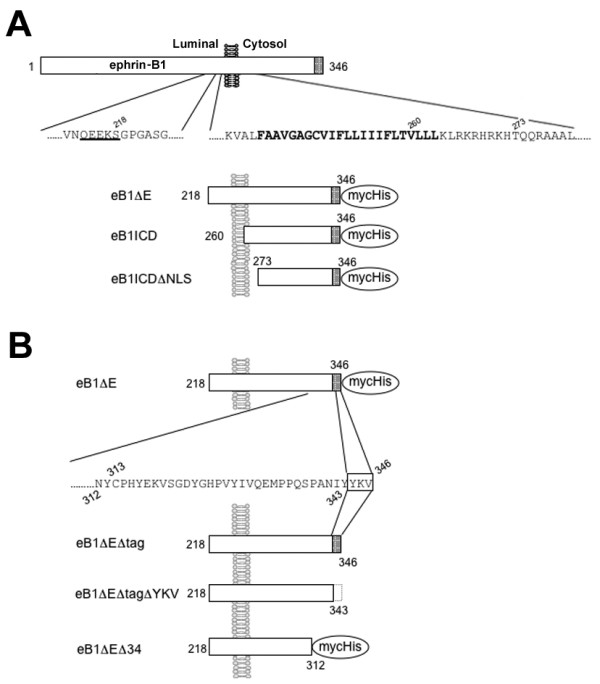
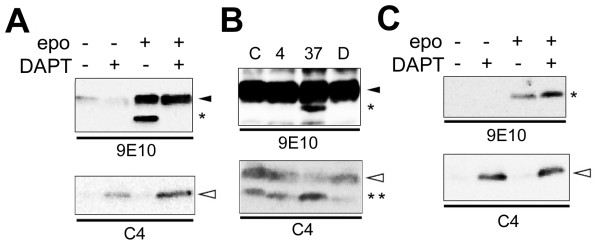
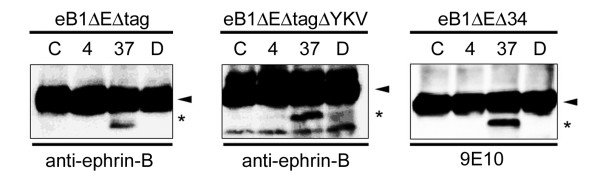
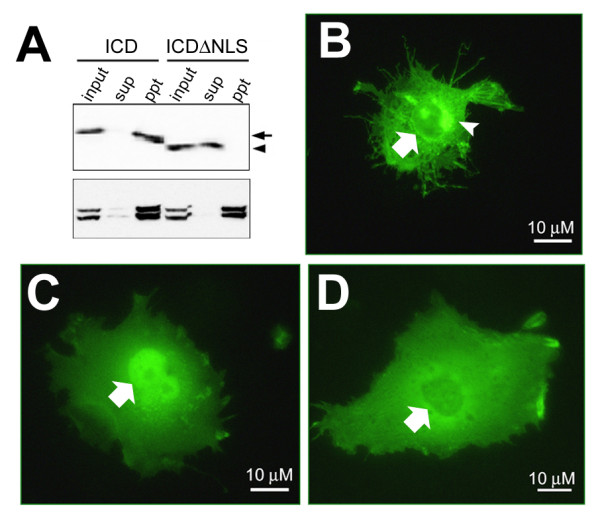
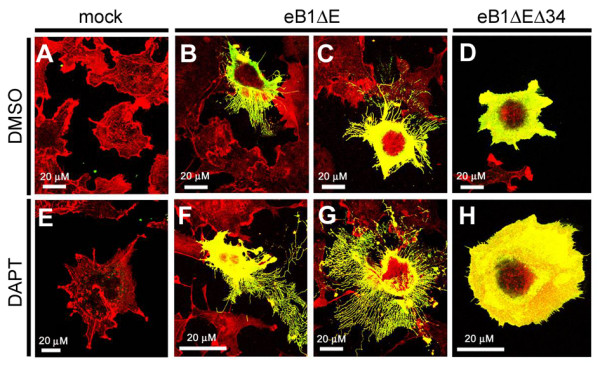
Results: We show that ephrin-B1, that participates in cell-cell repulsive and attractive signaling together with its Eph receptor, constitutively undergoes ectodomain shedding and that the residual membrane-tethered fragment is sequentially cleaved by gamma-secretase to release the intracellular domain. Furthermore, overexpression of membrane-tethered ephrin-B1 caused protrusion of numerous cellular processes consisted of F-actin, that required the preservation of the most C-terminal region of ephrin-B1. In contrast, soluble intracellular domain translocated into the nucleus and had no effect on cell morphology.
Conclusion: Our findings suggest that ephrin-B is a genuine substrate for gamma-secretase and regulates the cytoskeletal dynamics through intramembrane proteolysis.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
