

Casq2 deletion causes sarcoplasmic reticulum volume increase, premature Ca2+ release, and catecholaminergic polymorphic ventricular tachycardia
- PMID: 16932808
- PMCID: PMC1551934
- DOI: 10.1172/JCI29128
Casq2 deletion causes sarcoplasmic reticulum volume increase, premature Ca2+ release, and catecholaminergic polymorphic ventricular tachycardia
Abstract
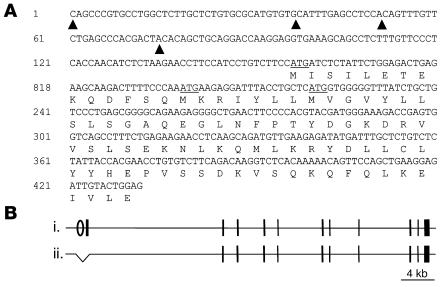
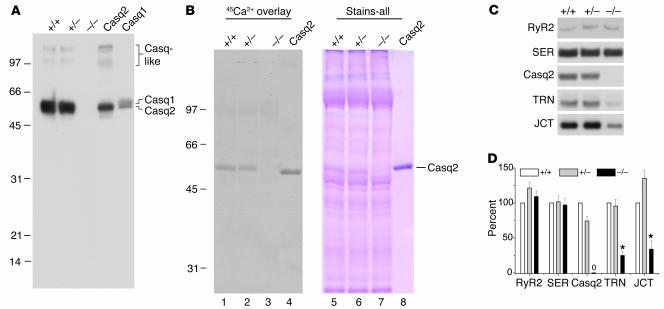
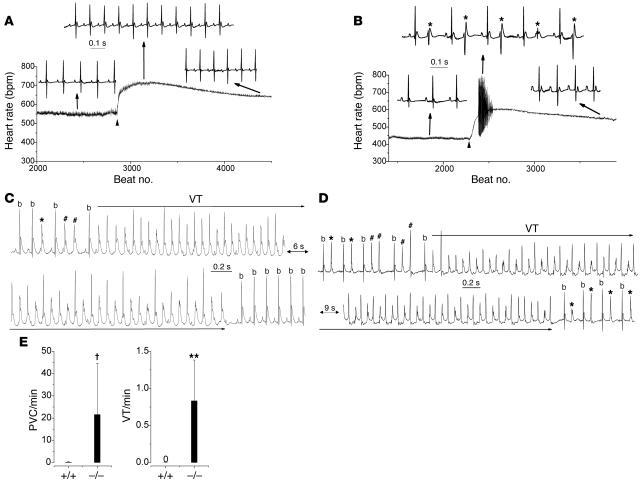
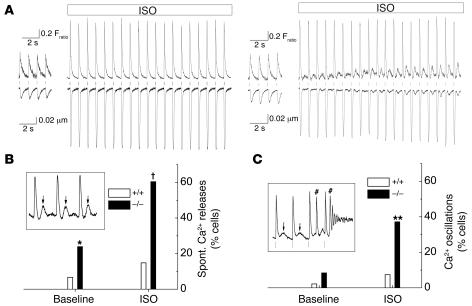
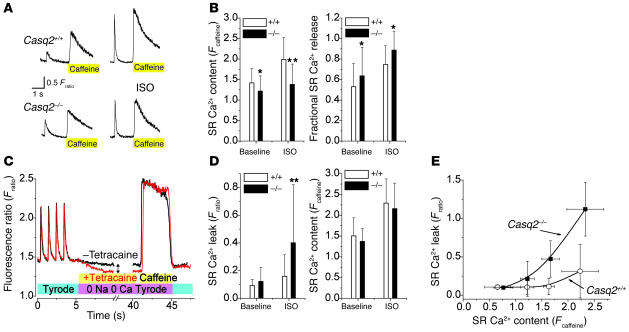
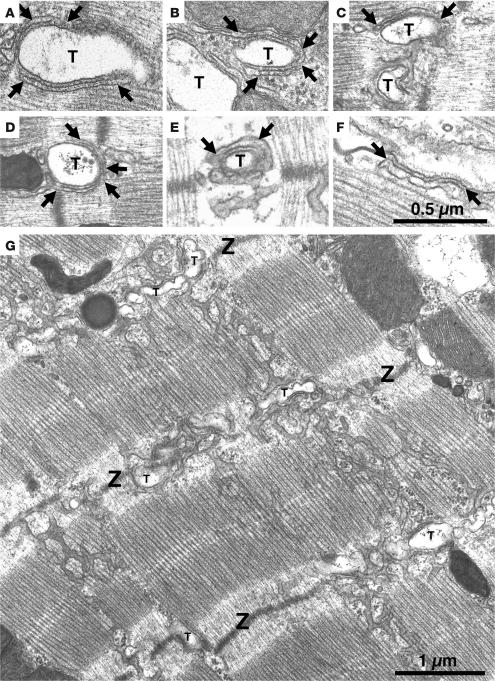
Cardiac calsequestrin (Casq2) is thought to be the key sarcoplasmic reticulum (SR) Ca2+ storage protein essential for SR Ca2+ release in mammalian heart. Human CASQ2 mutations are associated with catecholaminergic ventricular tachycardia. However, homozygous mutation carriers presumably lacking functional Casq2 display surprisingly normal cardiac contractility. Here we show that Casq2-null mice are viable and display normal SR Ca2+ release and contractile function under basal conditions. The mice exhibited striking increases in SR volume and near absence of the Casq2-binding proteins triadin-1 and junctin; upregulation of other Ca2+ -binding proteins was not apparent. Exposure to catecholamines in Casq2-null myocytes caused increased diastolic SR Ca2+ leak, resulting in premature spontaneous SR Ca2+ releases and triggered beats. In vivo, Casq2-null mice phenocopied the human arrhythmias. Thus, while the unique molecular and anatomic adaptive response to Casq2 deletion maintains functional SR Ca2+ storage, lack of Casq2 also causes increased diastolic SR Ca2+ leak, rendering Casq2-null mice susceptible to catecholaminergic ventricular arrhythmias.
Figures
References
-
- Scott B.T., Simmerman H.K., Collins J.H., Nadal-Ginard B., Jones L.R. Complete amino acid sequence of canine cardiac calsequestrin deduced by cDNA cloning. J. Biol. Chem. 1988;263:8958–8964. - PubMed
-
- Yano K., Zarain-Herzberg A. Sarcoplasmic reticulum calsequestrins: structural and functional properties. Mol. Cell. Biochem. 1994;135:61–70. - PubMed
-
- Cala S.E., Jones L.R. Rapid purification of calsequestrin from cardiac and skeletal muscle sarcoplasmic reticulum vesicles by Ca2+-dependent elution from phenyl-sepharose. J. Biol. Chem. 1983;258:11932–11936. - PubMed
-
- Campbell K.P., MacLennan D.H., Jorgensen A.O., Mintzer M.C. Purification and characterization of calsequestrin from canine cardiac sarcoplasmic reticulum and identification of the 53,000 dalton glycoprotein. J. Biol. Chem. 1983;258:1197–1204. - PubMed
-
- Jones L.R., Cala S.E. Biochemical evidence for functional heterogeneity of cardiac sarcoplasmic reticulum vesicles. J. Biol. Chem. 1981;256:11809–11818. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
