

The paradoxical pro- and anti-apoptotic actions of GSK3 in the intrinsic and extrinsic apoptosis signaling pathways
- PMID: 16935409
- PMCID: PMC1618798
- DOI: 10.1016/j.pneurobio.2006.07.006
The paradoxical pro- and anti-apoptotic actions of GSK3 in the intrinsic and extrinsic apoptosis signaling pathways
Abstract
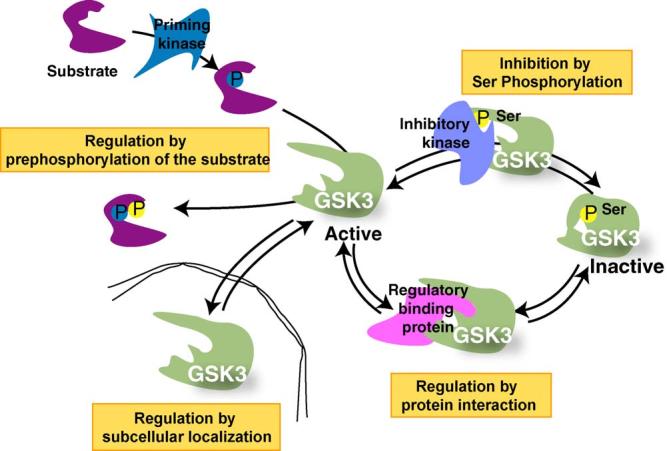
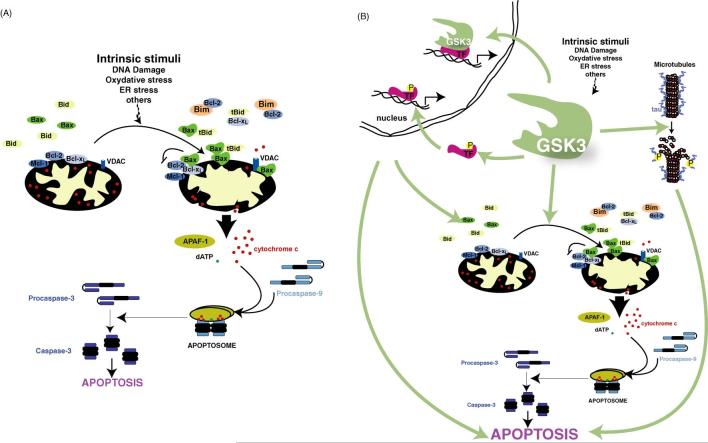
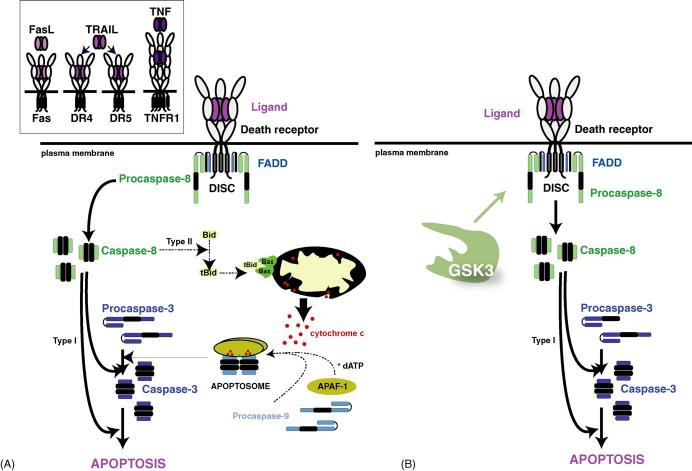
Few things can be considered to be more important to a cell than its threshold for apoptotic cell death, which can be modulated up or down, but rarely in both directions, by a single enzyme. Therefore, it came as quite a surprise to find that one enzyme, glycogen synthase kinase-3 (GSK3), has the perplexing capacity to either increase or decrease the apoptotic threshold. These apparently paradoxical effects now are known to be due to GSK3 oppositely regulating the two major apoptotic signaling pathways. GSK3 promotes cell death caused by the mitochondrial intrinsic apoptotic pathway, but inhibits the death receptor-mediated extrinsic apoptotic signaling pathway. Intrinsic apoptotic signaling, activated by cell damage, is promoted by GSK3 by facilitation of signals that cause disruption of mitochondria and by regulation of transcription factors that control the expression of anti- or pro-apoptotic proteins. The extrinsic apoptotic pathway entails extracellular ligands stimulating cell-surface death receptors that initiate apoptosis by activating caspase-8, and this early step in extrinsic apoptotic signaling is inhibited by GSK3. Thus, GSK3 modulates key steps in each of the two major pathways of apoptosis, but in opposite directions. Consequently, inhibitors of GSK3 provide protection from intrinsic apoptosis signaling but potentiate extrinsic apoptosis signaling. Studies of this eccentric ability of GSK3 to oppositely influence two types of apoptotic signaling have shed light on important regulatory mechanisms in apoptosis and provide the foundation for designing the rational use of GSK3 inhibitors for therapeutic interventions.
Figures
References
-
- Adhikary S, Eilers M. Transcriptional regulation and transformation by Myc proteins. Nat. Rev. Mol. Cell. Biol. 2005;6:635–645. - PubMed
-
- Akhtar RS, Ness JM, Roth KA. Bcl-2 family regulation of neuronal development and neurodegeneration. Biochim. Biophys. Acta. 2004;1644:189–203. - PubMed
-
- Alvarez G, Munoz-Montano JR, Satrustegui J, Avila J, Bogonez E, Diaz-Nido J. Lithium protects cultured neurons against β-amyloid-induced neurodegeneration. FEBS Lett. 1999;453:260–264. - PubMed
-
- Alvarez AR, Godoy JA, Mullendorff K, Olivares GH, Bronfman M, Inestrosa NC. Wnt-3a overcomes β-amyloid toxicity in rat hippocampal neurons. Exp. Cell Res. 2004;297:186–196. - PubMed
-
- Armstrong JS. Mitochondrial membrane permeabilization: the sine qua non for cell death. Bioessays. 2006;28:253–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
