

Rapid metabolism of glucose detected with FRET glucose nanosensors in epidermal cells and intact roots of Arabidopsis RNA-silencing mutants
- PMID: 16935985
- PMCID: PMC1560921
- DOI: 10.1105/tpc.106.044073
Rapid metabolism of glucose detected with FRET glucose nanosensors in epidermal cells and intact roots of Arabidopsis RNA-silencing mutants
Abstract
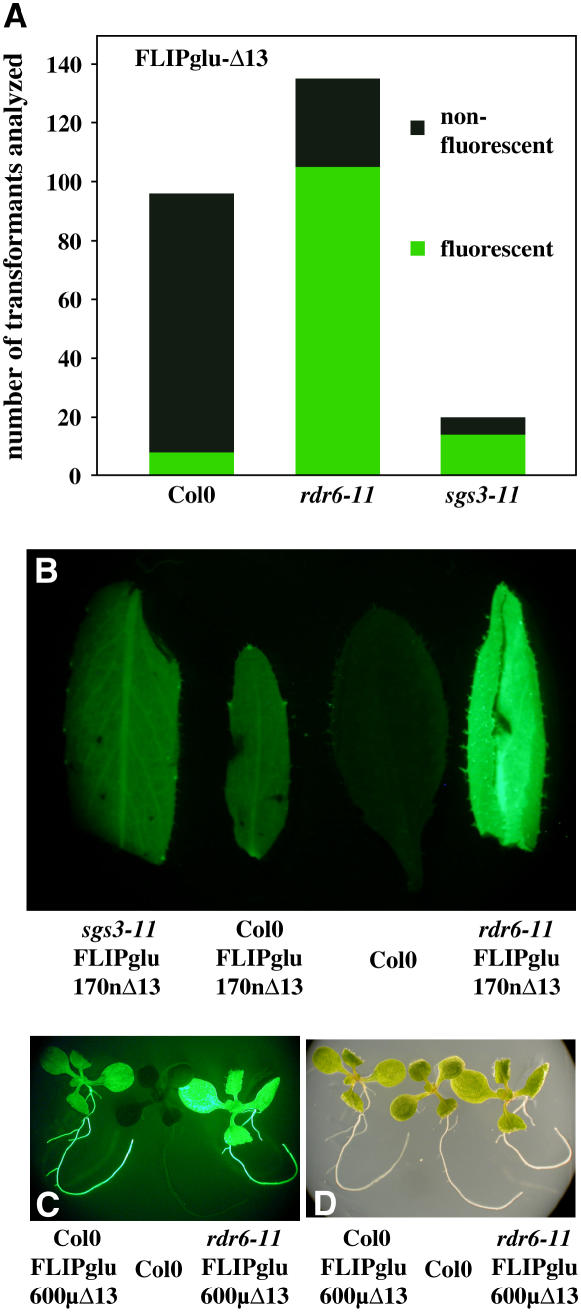
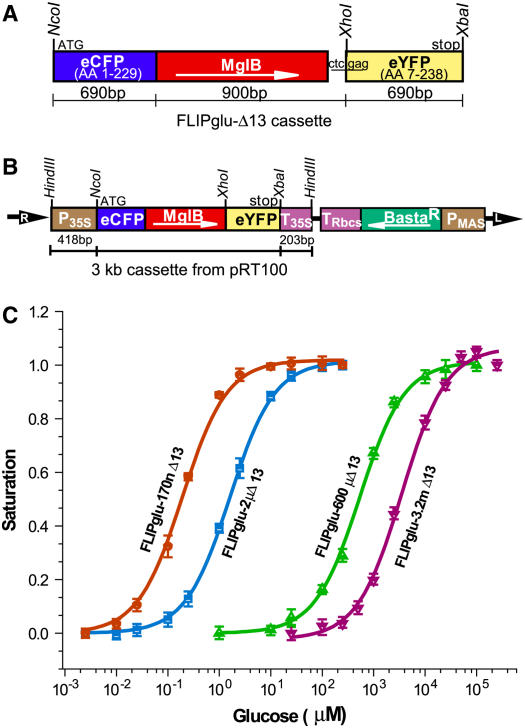
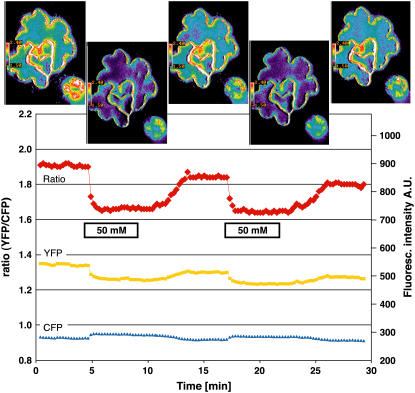
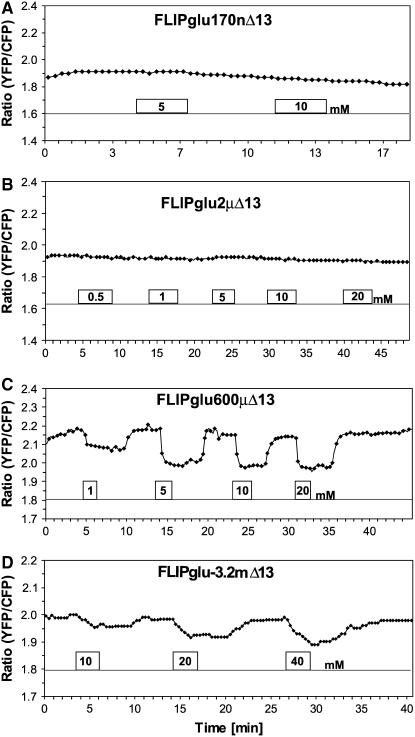
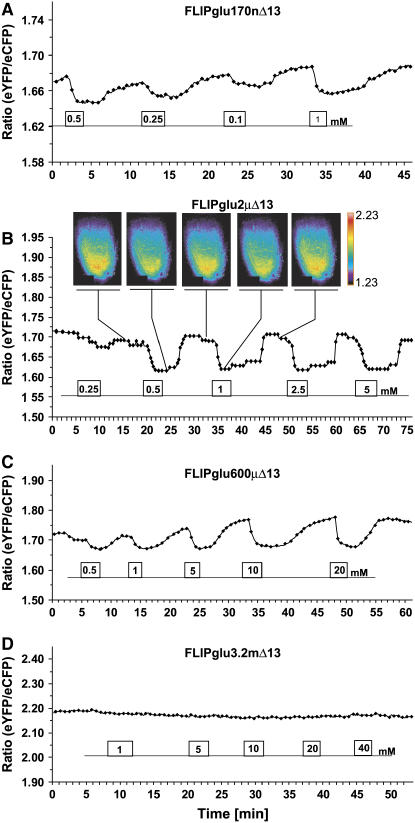
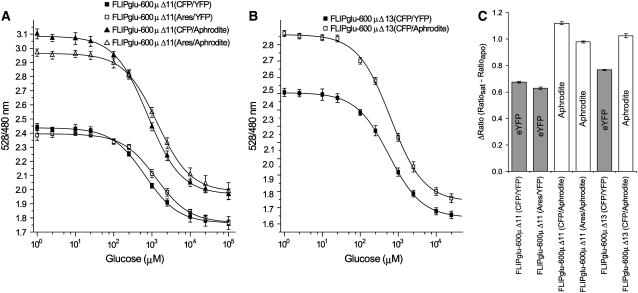
Genetically encoded glucose nanosensors have been used to measure steady state glucose levels in mammalian cytosol, nuclei, and endoplasmic reticulum. Unfortunately, the same nanosensors in Arabidopsis thaliana transformants manifested transgene silencing and undetectable fluorescence resonance energy transfer changes. Expressing nanosensors in sgs3 and rdr6 transgene silencing mutants eliminated silencing and resulted in high fluorescence levels. To measure glucose changes over a wide range (nanomolar to millimolar), nanosensors with higher signal-to-noise ratios were expressed in these mutants. Perfusion of leaf epidermis with glucose led to concentration-dependent ratio changes for nanosensors with in vitro K(d) values of 600 microM (FLIPglu-600 microDelta13) and 3.2 mM (FLIPglu-3.2 mDelta13), but one with 170 nM K(d) (FLIPglu-170 nDelta13) showed no response. In intact roots, FLIPglu-3.2 mDelta13 gave no response, whereas FLIPglu-600 microDelta13, FLIPglu-2 microDelta13, and FLIPglu-170 nDelta13 all responded to glucose. These results demonstrate that cytosolic steady state glucose levels depend on external supply in both leaves and roots, but under the conditions tested they are lower in root versus epidermal and guard cells. Without photosynthesis and external supply, cytosolic glucose can decrease to <90 nM in root cells. Thus, observed gradients are steeper than expected, and steady state levels do not appear subject to tight homeostatic control. Nanosensor-expressing plants can be used to assess glucose flux differences between cells, invertase-mediated sucrose hydrolysis in vivo, delivery of assimilates to roots, and glucose flux in mutants affected in sugar transport, metabolism, and signaling.
Figures
References
-
- Barros, L.F., Porras, O.H., and Bittner, C.X. (2005). Why glucose transport in the brain matters for PET. Trends Neurosci. 28 117–119. - PubMed
-
- Beclin, C., Boutet, S., Waterhouse, P., and Vaucheret, H. (2002). A branched pathway for transgene-induced RNA silencing in plants. Curr. Biol. 12 684–688. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
