

BAG3 deficiency results in fulminant myopathy and early lethality
- PMID: 16936253
- PMCID: PMC1698816
- DOI: 10.2353/ajpath.2006.060250
BAG3 deficiency results in fulminant myopathy and early lethality
Abstract
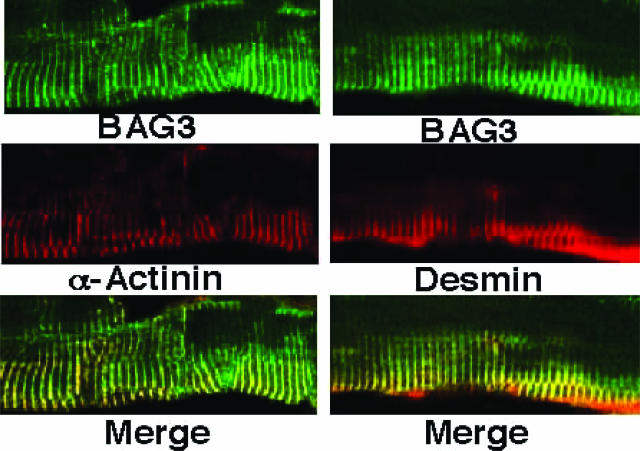
Bcl-2-associated athanogene 3 (BAG3) is a member of a conserved family of cyto-protective proteins that bind to and regulate Hsp70 family molecular chaperones. Here, we show that BAG3 is prominently expressed in striated muscle and colocalizes with Z-disks. Mice with homozygous disruption of the bag3 gene developed normally but deteriorated postnatally with stunted growth evident by 1 to 2 weeks of age and death by 4 weeks. BAG3-deficient animals developed a fulminant myopathy characterized by noninflammatory myofibrillar degeneration with apoptotic features. Knockdown of bag3 expression in cultured C2C12 myoblasts increased apoptosis on induction of differentiation, suggesting a need for bag3 for maintenance of myotube survival and confirming a cell autonomous role for bag3 in muscle. We conclude that although BAG3 is not required for muscle development, this co-chaperone appears to be critically important for maintenance of mature skeletal muscle.
Figures






References
-
- Takayama S, Xie Z, Reed J. An evolutionarily conserved family of Hsp70/Hsc70 molecular chaperone regulators. J Biol Chem. 1999;274:781–786. - PubMed
-
- Takayama S, Reed JC. Molecular chaperone targeting and regulation by BAG family proteins. Nature Cell Biol. 2001;3:E237–E241. - PubMed
-
- Takayama S, Sato T, Krajewski S, Kochel K, Irie S, Millan JA, Reed JC. Cloning and functional analysis of BAG-1: a novel Bcl-2 binding protein with anti-cell death activity. Cell. 1995;80:279–284. - PubMed
-
- Lee MY, Kim SY, Shin SL, Choi YS, Lee JH, Tsujimoto Y. Reactive astrocytes express bis, a bcl-2-binding protein, after transient forebrain ischemia. Exp Neurol. 2002;175:338–346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
