

Broad activation of the olfactory bulb produces long-lasting changes in odor perception
- PMID: 16938883
- PMCID: PMC1569199
- DOI: 10.1073/pnas.0602750103
Broad activation of the olfactory bulb produces long-lasting changes in odor perception
Abstract
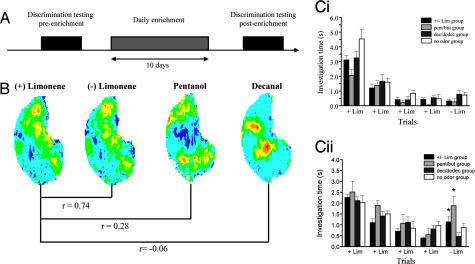
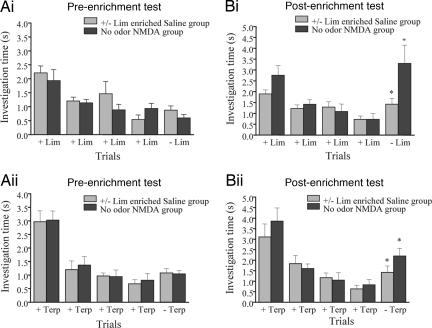
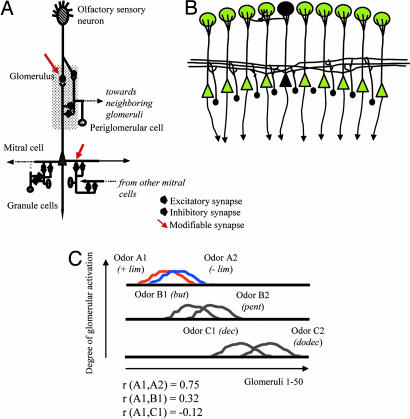
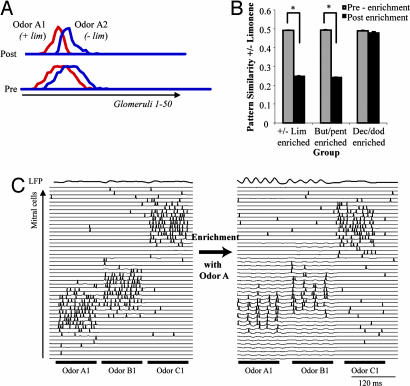
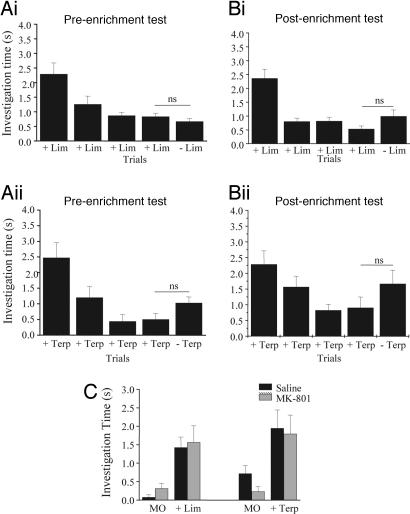
A number of electrophysiological experiments have shown that odor exposure alone, unaccompanied by behavioral training, changes the response patterns of neurons in the olfactory bulb. As a consequence of these changes, across mitral cells in the olfactory bulb, individual odors should be better discriminated because of previous exposure. We have previously shown that a daily 2-h exposure to odorants during 2 weeks enhances rats' ability to discriminate between chemically similar odorants. Here, we first show that the perception of test odorants is only modulated by enrichment with odorants that activate at least partially overlapping regions of the olfactory bulb. Second, we show that a broad activation of olfactory bulb neurons by daily local infusion of NMDA into both olfactory bulbs enhances the discrimination between chemically related odorants in a manner similar to the effect of daily exposure to odorants. Computational modeling of the olfactory bulb suggests that activity-dependent plasticity in the olfactory bulb can support the observed modulation in olfactory discrimination capability by enhancing contrast and synchronization in the olfactory bulb. Last, we show that blockade of NMDA receptors in the olfactory bulb impairs the effects of daily enrichment, suggesting that NMDA-dependent plasticity is involved in the changes in olfactory processing observed here.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Leggio M., Mandolesi L., Federico F., Spirito F., Ricci B., Gelfo F., Petrosini L. Behav. Brain Res. 2005;163:78–90. - PubMed
-
- Rampon C., Tang Y., Goodhouse J., Shimizu E., Kyin M., Tsien J. Nat. Neurosci. 2000;3:238–244. - PubMed
-
- Bennett J., McRae P., Levy L., Frick K. Neurobiol. Learn. Mem. 2005;85:139–152. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
