

Plasticity of functional connectivity in the adult spinal cord
- PMID: 16939979
- PMCID: PMC1664672
- DOI: 10.1098/rstb.2006.1884
Plasticity of functional connectivity in the adult spinal cord
Abstract
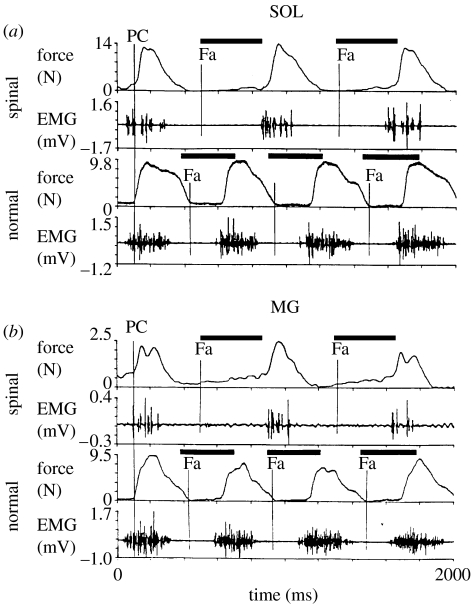
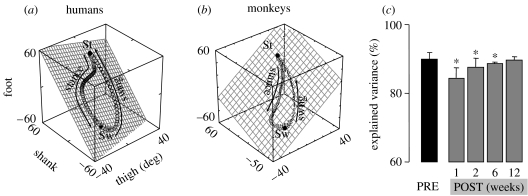
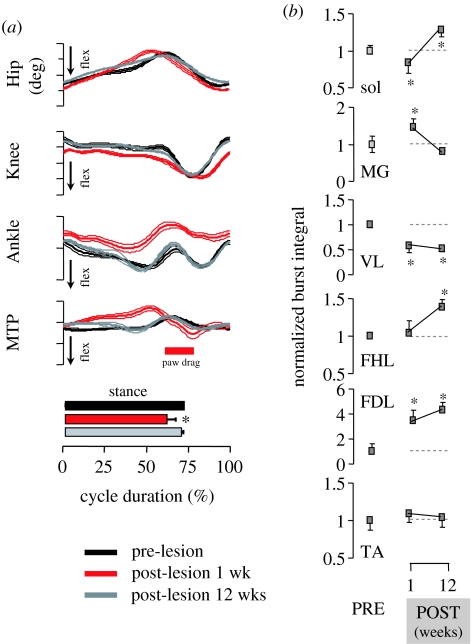
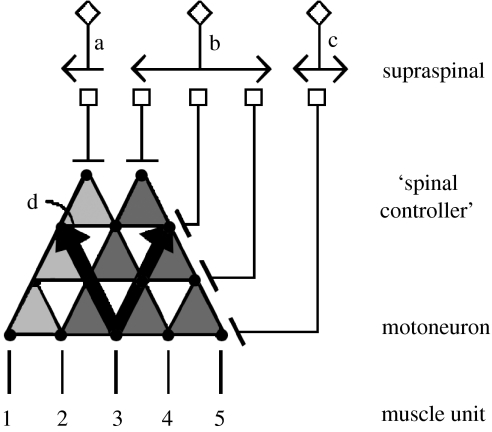
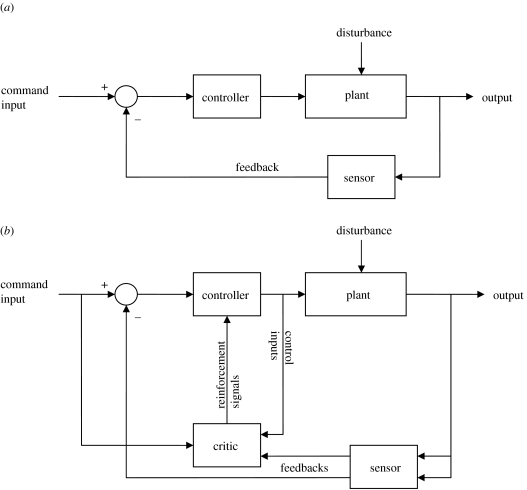
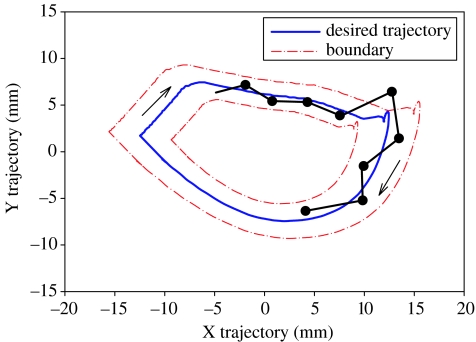
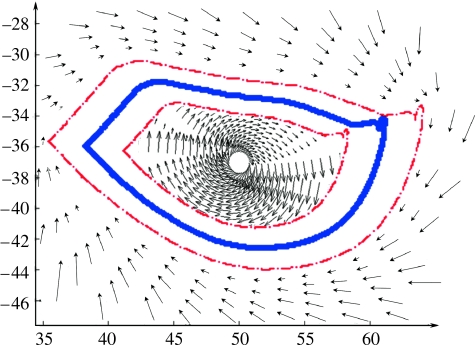
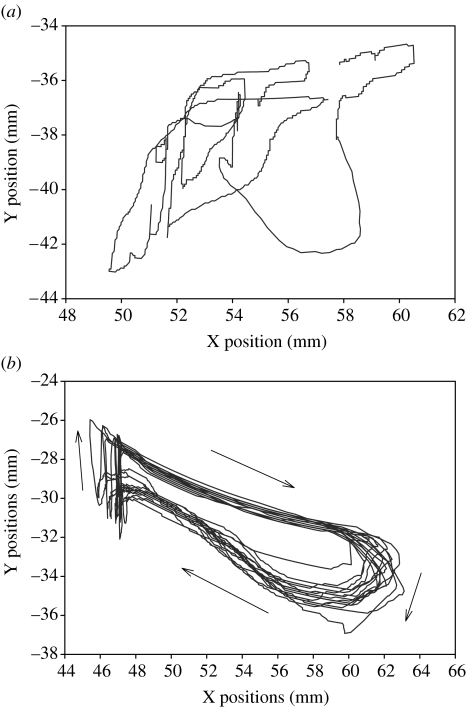
This paper emphasizes several characteristics of the neural control of locomotion that provide opportunities for developing strategies to maximize the recovery of postural and locomotor functions after a spinal cord injury (SCI). The major points of this paper are: (i) the circuitry that controls standing and stepping is extremely malleable and reflects a continuously varying combination of neurons that are activated when executing stereotypical movements; (ii) the connectivity between neurons is more accurately perceived as a functional rather than as an anatomical phenomenon; (iii) the functional connectivity that controls standing and stepping reflects the physiological state of a given assembly of synapses, where the probability of these synaptic events is not deterministic; (iv) rather, this probability can be modulated by other factors such as pharmacological agents, epidural stimulation and/or motor training; (v) the variability observed in the kinematics of consecutive steps reflects a fundamental feature of the neural control system and (vi) machine-learning theories elucidate the need to accommodate variability in developing strategies designed to enhance motor performance by motor training using robotic devices after an SCI.
Figures
References
-
- Anderson J.R, Michalski R.S, Carbonell J.G, Mitchell T.M. M. Kaufmann; Los Altos, CA: 1983. Machine learning: an artificial intelligence approach.
-
- Barbeau H, Fung J, Leroux A, Ladouceur M. A review of the adaptability and recovery of locomotion after spinal cord injury. Prog. Brain Res. 2002;137:9–25. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical