

Molecular architecture of a eukaryotic DNA replication terminus-terminator protein complex
- PMID: 16940176
- PMCID: PMC1636744
- DOI: 10.1128/MCB.01102-06
Molecular architecture of a eukaryotic DNA replication terminus-terminator protein complex
Abstract
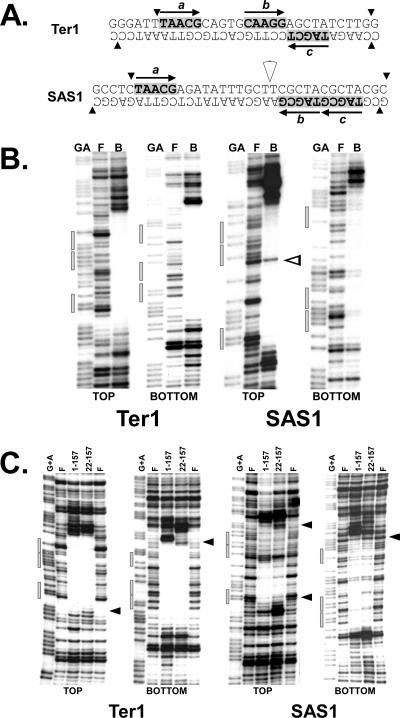
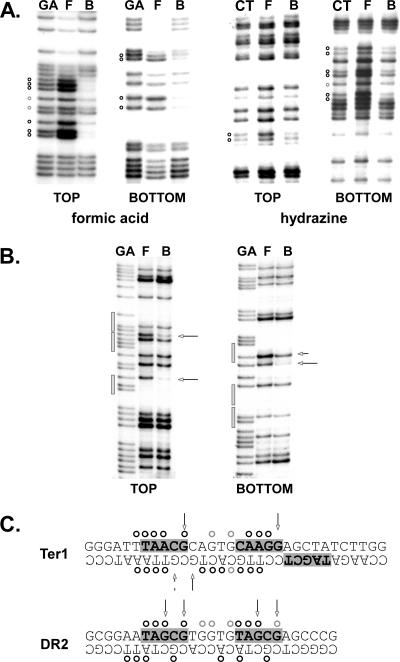
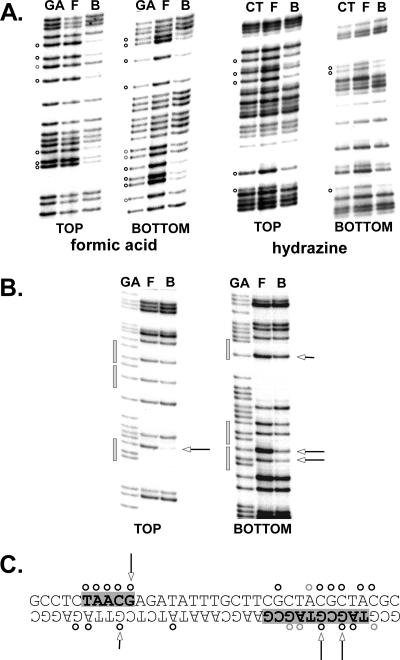
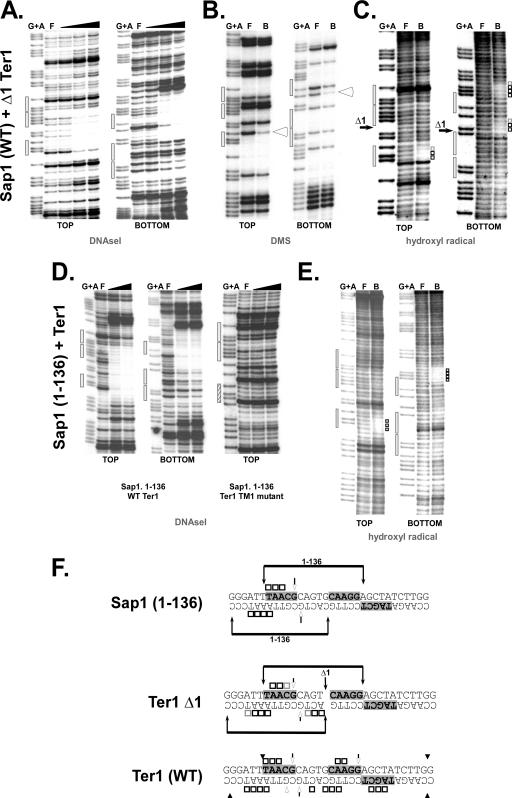
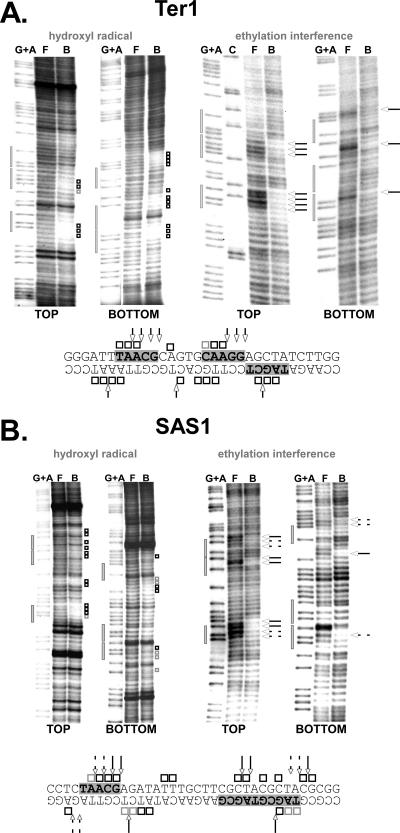
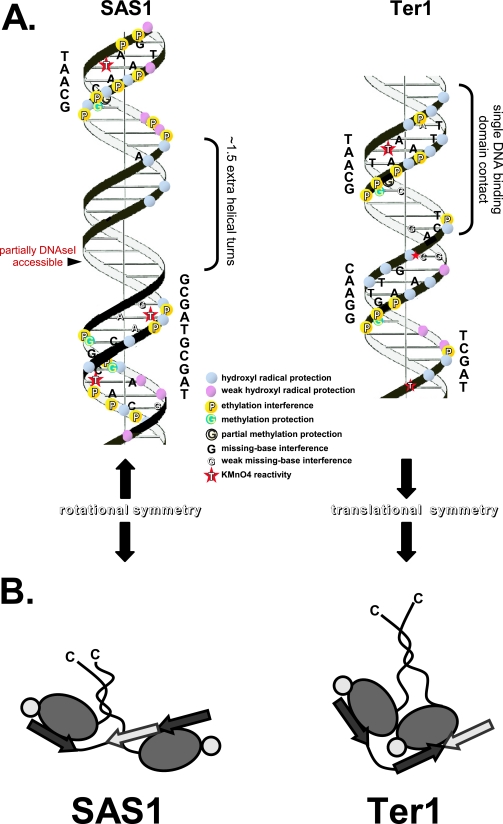
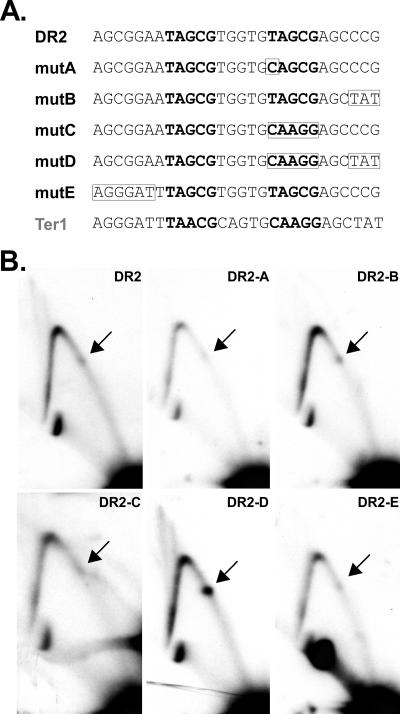
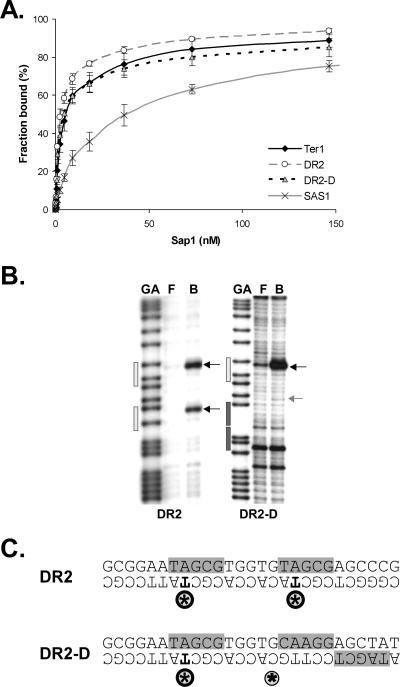
DNA replication forks pause at programmed fork barriers within nontranscribed regions of the ribosomal DNA (rDNA) genes of many eukaryotes to coordinate and regulate replication, transcription, and recombination. The mechanism of eukaryotic fork arrest remains unknown. In Schizosaccharomyces pombe, the promiscuous DNA binding protein Sap1 not only causes polar fork arrest at the rDNA fork barrier Ter1 but also regulates mat1 imprinting at SAS1 without fork pausing. Towards an understanding of eukaryotic fork arrest, we probed the interactions of Sap1 with Ter1 as contrasted with SAS1. The Sap1 dimer bound Ter1 with high affinity at one face of the DNA, contacting successive major grooves. The complex displayed translational symmetry. In contrast, Sap1 subunits approached SAS1 from opposite helical faces, forming a low-affinity complex with mirror image rotational symmetry. The alternate symmetries were reflected in distinct Sap1-induced helical distortions. Importantly, modulating protein-DNA interactions of the fork-proximal Sap1 subunit with the nonnatural binding site DR2 affected blocking efficiency without changes in binding affinity or binding mode but with alterations in Sap1-induced DNA distortion. The results reveal that Sap1-DNA affinity alone is insufficient to account for fork arrest and suggest that Sap1 binding-induced structural changes may result in formation of a competent fork-blocking complex.
Figures

Similar articles
-
Sap1p binds to Ter1 at the ribosomal DNA of Schizosaccharomyces pombe and causes polar replication fork arrest.J Biol Chem. 2005 Nov 25;280(47):39135-42. doi: 10.1074/jbc.M508996200. Epub 2005 Sep 28. J Biol Chem. 2005. PMID: 16195226
-
Structure of the replication regulator Sap1 reveals functionally important interfaces.Sci Rep. 2018 Jul 19;8(1):10930. doi: 10.1038/s41598-018-29198-9. Sci Rep. 2018. PMID: 30026545 Free PMC article.
-
The mating type switch-activating protein Sap1 Is required for replication fork arrest at the rRNA genes of fission yeast.Mol Cell Biol. 2005 Oct;25(19):8755-61. doi: 10.1128/MCB.25.19.8755-8761.2005. Mol Cell Biol. 2005. PMID: 16166653 Free PMC article.
-
Mechanisms of polar arrest of a replication fork.Mol Microbiol. 2009 Apr;72(2):279-85. doi: 10.1111/j.1365-2958.2009.06656.x. Epub 2009 Mar 4. Mol Microbiol. 2009. PMID: 19298368 Review.
-
Evolutionary diversification of eukaryotic DNA replication machinery.Subcell Biochem. 2012;62:19-35. doi: 10.1007/978-94-007-4572-8_2. Subcell Biochem. 2012. PMID: 22918578 Review.
Cited by
-
Replication fork blocking deficiency leads to a reduction of rDNA copy number in budding yeast.iScience. 2024 Feb 6;27(3):109120. doi: 10.1016/j.isci.2024.109120. eCollection 2024 Mar 15. iScience. 2024. PMID: 38384843 Free PMC article.
-
The Fission Yeast Mating-Type Switching Motto: "One-for-Two" and "Two-for-One".Microbiol Mol Biol Rev. 2023 Mar 21;87(1):e0000821. doi: 10.1128/mmbr.00008-21. Epub 2023 Jan 11. Microbiol Mol Biol Rev. 2023. PMID: 36629411 Free PMC article. Review.
-
Mechanistic insights into replication termination as revealed by investigations of the Reb1-Ter3 complex of Schizosaccharomyces pombe.Mol Cell Biol. 2008 Nov;28(22):6844-57. doi: 10.1128/MCB.01235-08. Epub 2008 Sep 15. Mol Cell Biol. 2008. PMID: 18794373 Free PMC article.
-
Repeat expansion in the budding yeast ribosomal DNA can occur independently of the canonical homologous recombination machinery.Nucleic Acids Res. 2011 Nov 1;39(20):8778-91. doi: 10.1093/nar/gkr589. Epub 2011 Jul 17. Nucleic Acids Res. 2011. PMID: 21768125 Free PMC article.
-
Phosphorylation of CMG helicase and Tof1 is required for programmed fork arrest.Proc Natl Acad Sci U S A. 2016 Jun 28;113(26):E3639-48. doi: 10.1073/pnas.1607552113. Epub 2016 Jun 13. Proc Natl Acad Sci U S A. 2016. PMID: 27298353 Free PMC article.
References
-
- Bada, M., D. Walther, B. Arcangioli, S. Doniach, and M. Delarue. 2000. Solution structural studies and low-resolution model of the Schizosaccharomyces pombe sap1 protein. J. Mol. Biol. 300:563-574. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources