

Gata4 is essential for the maintenance of jejunal-ileal identities in the adult mouse small intestine
- PMID: 16940177
- PMCID: PMC1636804
- DOI: 10.1128/MCB.00124-06
Gata4 is essential for the maintenance of jejunal-ileal identities in the adult mouse small intestine
Abstract
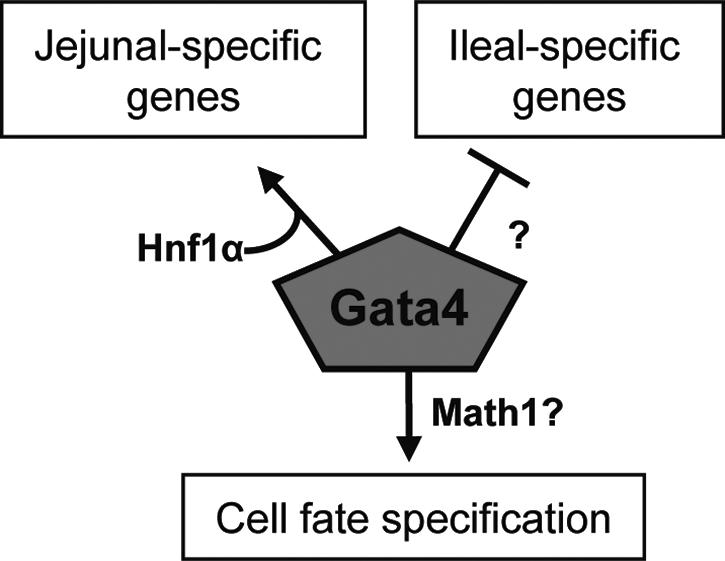
Gata4, a member of the zinc finger family of GATA transcription factors, is highly expressed in duodenum and jejunum but is nearly undetectable in distal ileum of adult mice. We show here that the caudal reduction of Gata4 is conserved in humans. To test the hypothesis that the regional expression of Gata4 is critical for the maintenance of jejunal-ileal homeostasis in the adult small intestine in vivo, we established an inducible, intestine-specific model that results in the synthesis of a transcriptionally inactive Gata4 mutant. Synthesis of mutant Gata4 in jejuna of 6- to 8-week-old mice resulted in an attenuation of absorptive enterocyte genes normally expressed in jejunum but not in ileum, including those for the anticipated targets liver fatty acid binding protein (Fabp1) and lactase-phlorizin hydrolase (LPH), and a surprising induction of genes normally silent in jejunum but highly expressed in ileum, specifically those involved in bile acid transport. Inactivation of Gata4 resulted in an increase in the goblet cell population and a redistribution of the enteroendocrine subpopulations, all toward an ileal phenotype. The gene encoding Math1, a known activator of the secretory cell fate, was induced approximately 75% (P < 0.05). Gata4 is thus an important positional signal required for the maintenance of jejunal-ileal identities in the adult mouse small intestine.
Figures
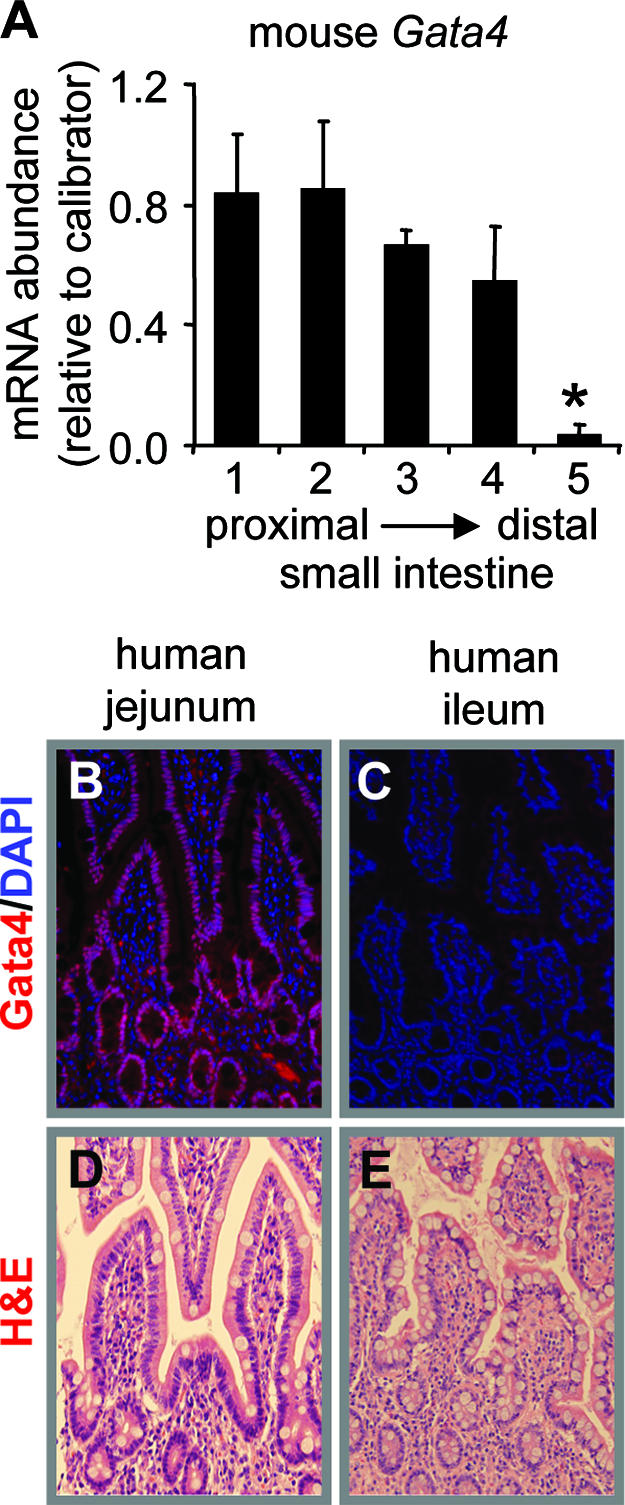
 , P < 0.05; mean ± SEM, n = 3). The calibrator was a pooled sample of jejunal RNA. (B and C) Immunofluorescence of adult human intestinal epithelium for Gata4 (red) and nucleic acid by DAPI (blue), showing the presence of Gata4 in jejunum (B) and the absence of Gata4 in ileum (C). (D and E). H&E staining of adult human jejunum and ileum, showing intact morphology.
, P < 0.05; mean ± SEM, n = 3). The calibrator was a pooled sample of jejunal RNA. (B and C) Immunofluorescence of adult human intestinal epithelium for Gata4 (red) and nucleic acid by DAPI (blue), showing the presence of Gata4 in jejunum (B) and the absence of Gata4 in ileum (C). (D and E). H&E staining of adult human jejunum and ileum, showing intact morphology.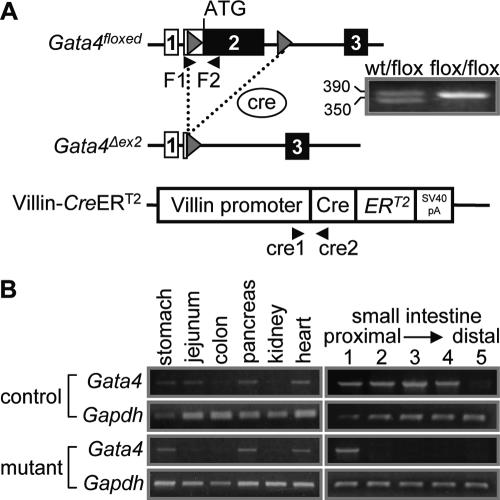
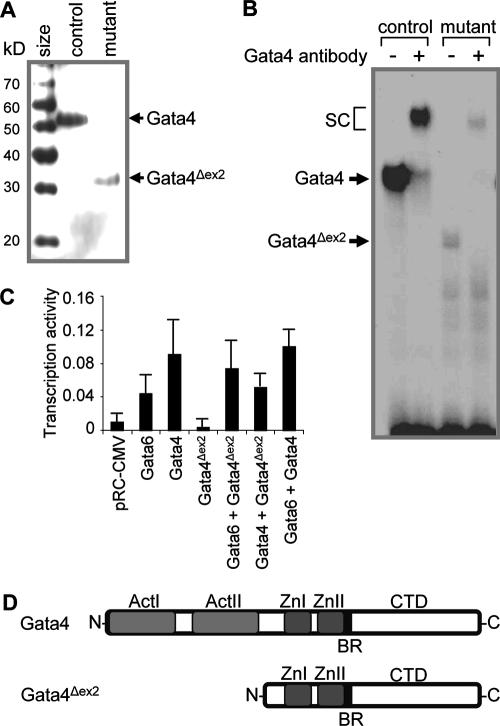
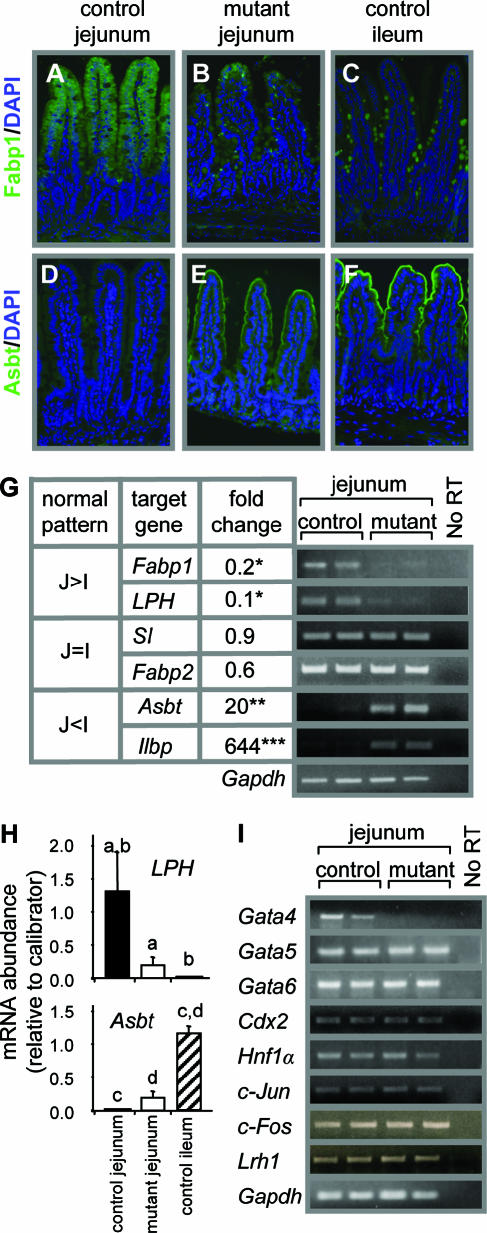 , P < 0.05; , P < 0.01; , P < 0.001) of genes normally expressed at higher levels in jejunum than in ileum (J>I), equally in jejunum and ileum (J=I), and at lower levels in jejunum than in ileum (J<I). Semiquantitative RT-PCR (right) is shown for two representative samples each from control and Gata4 mutant mice. A reaction without reverse transcriptase (No RT) served as a control for DNA contamination. (H) Comparison of LPH and Asbt mRNAs in control jejunum, mutant jejunum, and control ileum by real-time RT-PCR, showing that the transformation to an ileal-like phenotype is not complete. Data are means ± SEMs (n = 5). Bars with the same letter are significantly different from each other (P < 0.05). The calibrators were adult jejunal RNA for LPH and adult ileal RNA for Asbt. (I) Semiquantitative RT-PCR analysis for Gata5, Gata6, Cdx2, Hnf1α, c-Jun, c-Fos, Lrh1, and Gapdh on RNA from jejunum, showing that the mRNA abundances in control and Gata4 mutant mice are not different. A reaction without reverse transcriptase (No RT) served as a control for DNA contamination.
, P < 0.05; , P < 0.01; , P < 0.001) of genes normally expressed at higher levels in jejunum than in ileum (J>I), equally in jejunum and ileum (J=I), and at lower levels in jejunum than in ileum (J<I). Semiquantitative RT-PCR (right) is shown for two representative samples each from control and Gata4 mutant mice. A reaction without reverse transcriptase (No RT) served as a control for DNA contamination. (H) Comparison of LPH and Asbt mRNAs in control jejunum, mutant jejunum, and control ileum by real-time RT-PCR, showing that the transformation to an ileal-like phenotype is not complete. Data are means ± SEMs (n = 5). Bars with the same letter are significantly different from each other (P < 0.05). The calibrators were adult jejunal RNA for LPH and adult ileal RNA for Asbt. (I) Semiquantitative RT-PCR analysis for Gata5, Gata6, Cdx2, Hnf1α, c-Jun, c-Fos, Lrh1, and Gapdh on RNA from jejunum, showing that the mRNA abundances in control and Gata4 mutant mice are not different. A reaction without reverse transcriptase (No RT) served as a control for DNA contamination.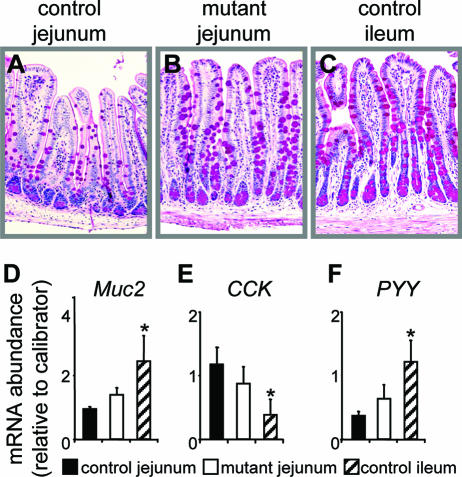 , P < 0.05 compared to control jejunum; data are means ± SEMs.
, P < 0.05 compared to control jejunum; data are means ± SEMs.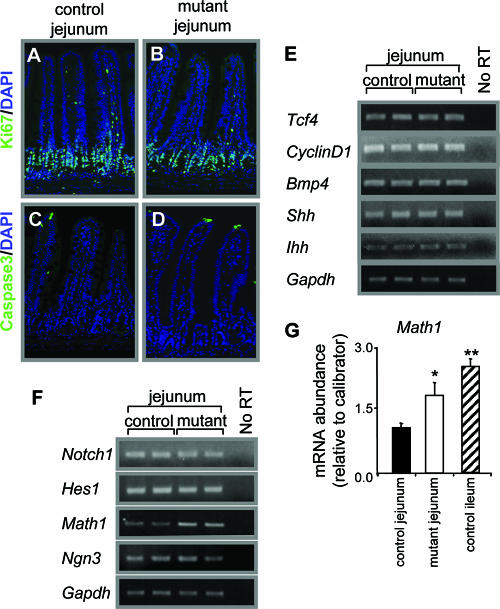
References
-
- Bjerknes, M., and H. Chang. 2005. Gastrointestinal stem cells. II. Intestinal stem cells. Am. J. Physiol. 289:G381-G387. - PubMed
-
- Bosse, T., H. M. van Wering, M. Gielen, L. N. Dowling, J. J. Fialkovich, C. M. Piaseckyj, F. J. Gonzalez, T. E. Akiyama, R. K. Montgomery, R. J. Grand, and S. D. Krasinski. 2006. Hepatocyte nuclear factor-1α is required for expression, but dispensable for histone acetylation of the lactase-phlorizin hydrolase gene in vivo. Am. J. Physiol. 290:1016-1024. - PubMed
-
- Boudreau, F., E. H. Rings, H. M. van Wering, R. K. Kim, G. P. Swain, S. D. Krasinski, J. Moffett, R. J. Grand, E. R. Suh, and P. G. Traber. 2002. Hepatocyte nuclear factor-1α, GATA-4, and caudal related homeodomain protein Cdx2 interact funtionally to modulate intestinal gene transcription. Implications for the developmental regulation of the sucrase-isomaltase gene. J. Biol. Chem. 277:31909-31917. - PubMed
-
- Chen, F., L. Ma, N. Al-Ansari, and B. L. Shneider. 2001. The role of AP-1 in the transcriptional regulation of the rat apical sodium-dependent bile acid transporter. J. Biol. Chem. 276:38703-38714. - PubMed
-
- Chen, F., L. Ma, P. A. Dawson, C. J. Sinal, E. Sehayek, F. J. Gonzalez, J. L. Breslow, M. Ananthanarayanan, and B. L. Shneider. 2003. Liver receptor homologue-1 mediates species- and cell line-specific bile acid-dependent negative feedback regulation of the apical sodium-dependent bile acid transporter. J. Biol. Chem. 278:19909-19916. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases