

Mutation at residue 523 creates a second receptor binding site on human parainfluenza virus type 1 hemagglutinin-neuraminidase protein
- PMID: 16940513
- PMCID: PMC1563932
- DOI: 10.1128/JVI.00969-06
Mutation at residue 523 creates a second receptor binding site on human parainfluenza virus type 1 hemagglutinin-neuraminidase protein
Abstract
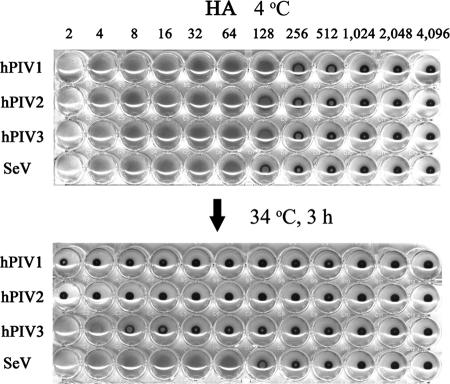
The paramyxovirus hemagglutinin-neuraminidase (HN) is a multifunctional protein mediating hemagglutination (HA), neuraminidase (NA), and fusion promotion activities. It has been a matter of debate whether HN contains combined or separate sites for HA and NA activities. To clear the issue, we determined the presence of the second binding site on human parainfluenza virus (hPIV) type 1, 2, and 3 and Sendai virus (SeV) HN proteins. Results of virus elution from erythrocytes at an elevated temperature and HA inhibition by NA inhibitor BCX-2798 suggest that all hPIVs bind to the receptor only through the NA catalytic site, while SeV HN has an additional receptor binding site. Comparison of SeV and hPIV1 HN sequences revealed two amino acid differences at residues 521 and 523 in the region close to the second binding site identified in Newcastle disease virus HN. We mutated hPIV1 HN at position 523 from Asn to the residue of SeV HN, Asp, and rescued a recombinant SeV that carries the mutated hPIV1 HN by a reverse genetics system. The hPIV1 HN with Asp at position 523 hemagglutinated in the presence of BCX-2798, suggesting that the amino acid difference at position 523 is critical for the formation of a second binding site. Creation of the second binding site on hPIV1 HN, however, did not significantly affect the growth or fusion activity of the recombinant virus. Our study indicates that the presence and requirement of a second binding site vary among paramyxoviruses.
Figures
References
-
- Alymova, I. V., G. Taylor, T. Takimoto, T. H. Lin, P. Chand, Y. S. Babu, C. Li, X. Xiong, and A. Portner. 2004. Efficacy of novel hemagglutinin-neuraminidase inhibitors BCX 2798 and BCX 2855 against human parainfluenza viruses in vitro and in vivo. Antimicrob. Agents Chemother. 48:1495-1502. - PMC - PubMed
-
- Bousse, T., T. Takimoto, W. L. Gorman, T. Takahashi, and A. Portner. 1994. Regions on the hemagglutinin-neuraminidase proteins of human parainfluenza virus type-1 and Sendai virus important for membrane fusion. Virology 204:506-514. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
