

Functional requirement of noncoding Y RNAs for human chromosomal DNA replication
- PMID: 16943439
- PMCID: PMC1592862
- DOI: 10.1128/MCB.01060-06
Functional requirement of noncoding Y RNAs for human chromosomal DNA replication
Abstract
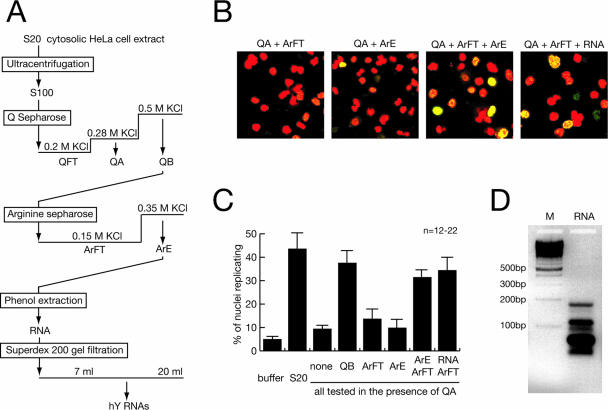
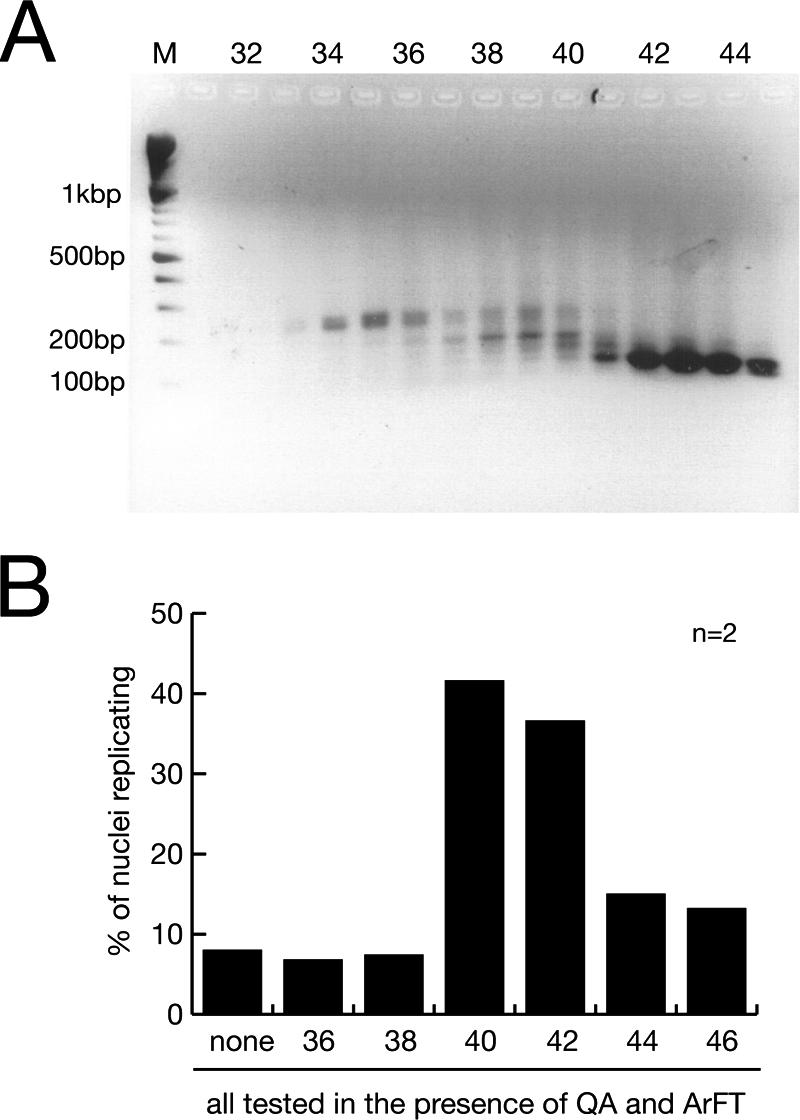
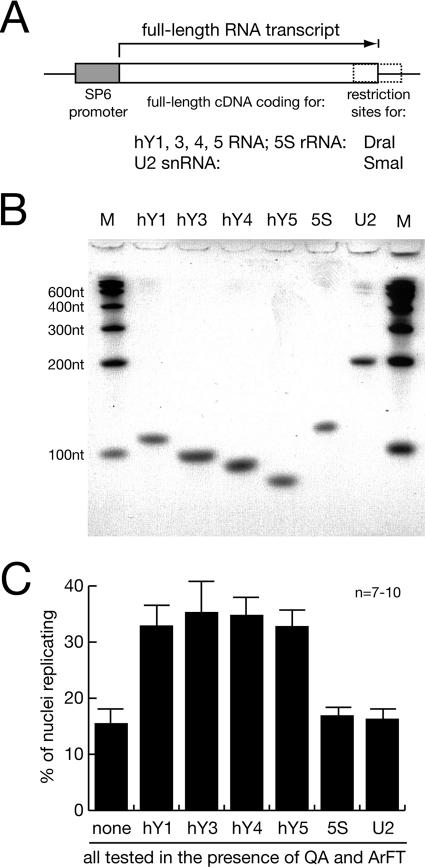
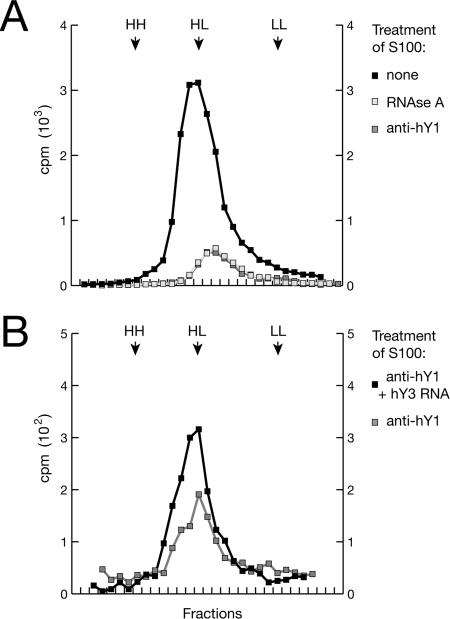
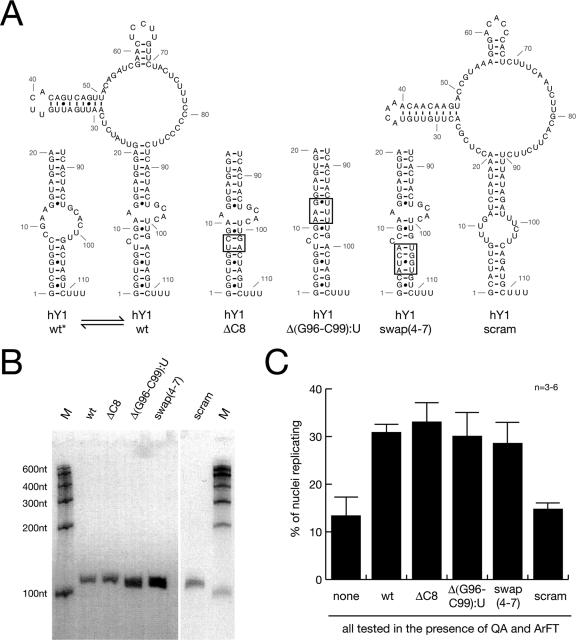
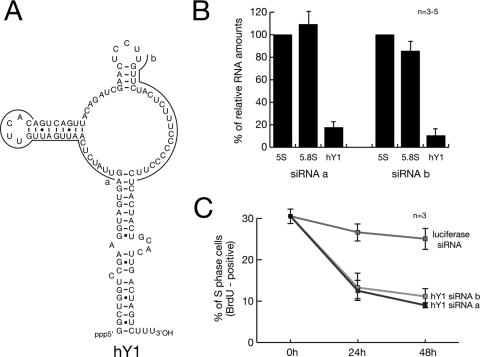
Noncoding RNAs are recognized increasingly as important regulators of fundamental biological processes, such as gene expression and development, in eukaryotes. We report here the identification and functional characterization of the small noncoding human Y RNAs (hY RNAs) as novel factors for chromosomal DNA replication in a human cell-free system. In addition to protein fractions, hY RNAs are essential for the establishment of active chromosomal DNA replication forks in template nuclei isolated from late-G(1)-phase human cells. Specific degradation of hY RNAs leads to the inhibition of semiconservative DNA replication in late-G(1)-phase template nuclei. This inhibition is negated by resupplementation of hY RNAs. All four hY RNAs (hY1, hY3, hY4, and hY5) can functionally substitute for each other in this system. Mutagenesis of hY1 RNA showed that the binding site for Ro60 protein, which is required for Ro RNP assembly, is not essential for DNA replication. Degradation of hY1 RNA in asynchronously proliferating HeLa cells by RNA interference reduced the percentages of cells incorporating bromodeoxyuridine in vivo. These experiments implicate a functional role for hY RNAs in human chromosomal DNA replication.
Figures


References
-
- Arias, E. E., and J. C. Walter. 2004. Initiation of DNA replication in xenopus egg extracts. Front. Biosci. 9:3029-3045. - PubMed
-
- Bernstein, E., and C. D. Allis. 2005. RNA meets chromatin. Genes Dev. 19:1635-1655. - PubMed
-
- Chen, X., and S. L. Wolin. 2004. The Ro 60 kDa autoantigen: insights into cellular function and role in autoimmunity. J. Mol. Med. 82:232-239. - PubMed
-
- Coverley, D., H. Laman, and R. A. Laskey. 2002. Distinct roles for cyclins E and A during DNA replication complex assembly and activation. Nat. Cell Biol. 4:523-528. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials