

Activation of pedunculopontine tegmental protein kinase A: a mechanism for rapid eye movement sleep generation in the freely moving rat
- PMID: 16943549
- PMCID: PMC6675344
- DOI: 10.1523/JNEUROSCI.2173-06.2006
Activation of pedunculopontine tegmental protein kinase A: a mechanism for rapid eye movement sleep generation in the freely moving rat
Abstract
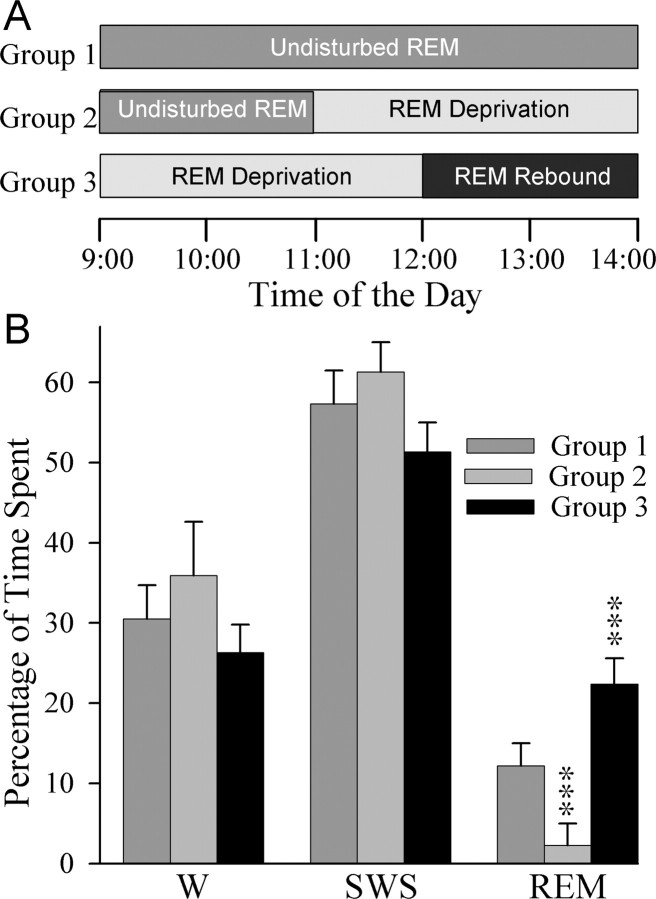
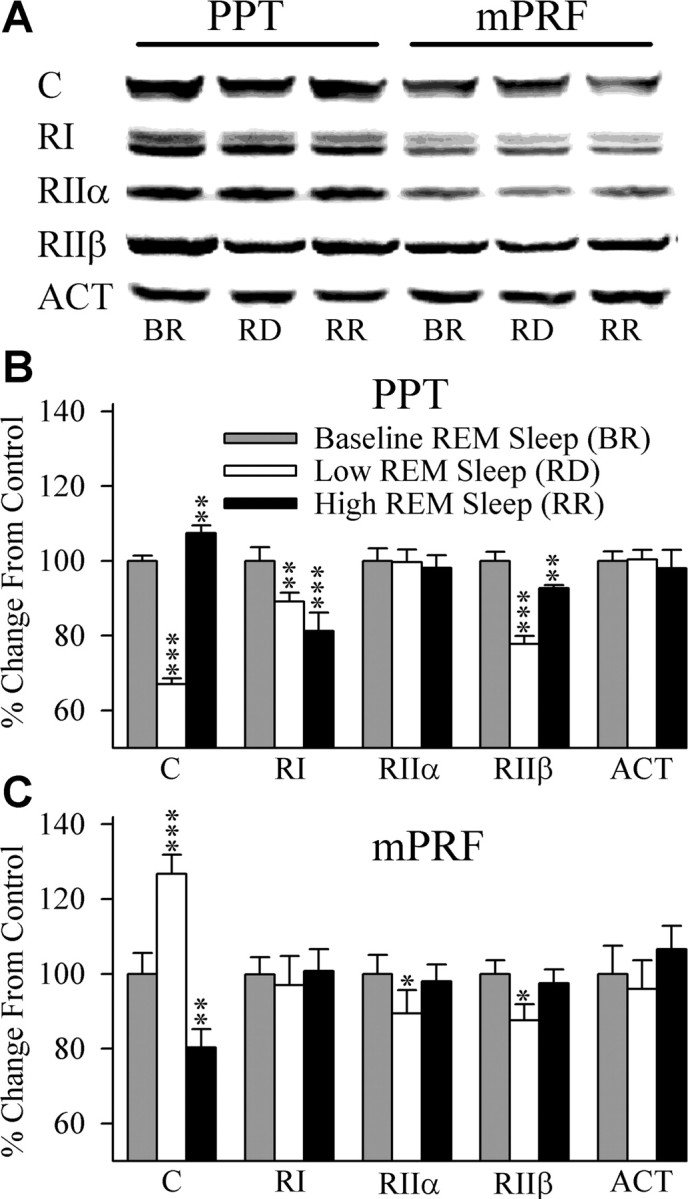
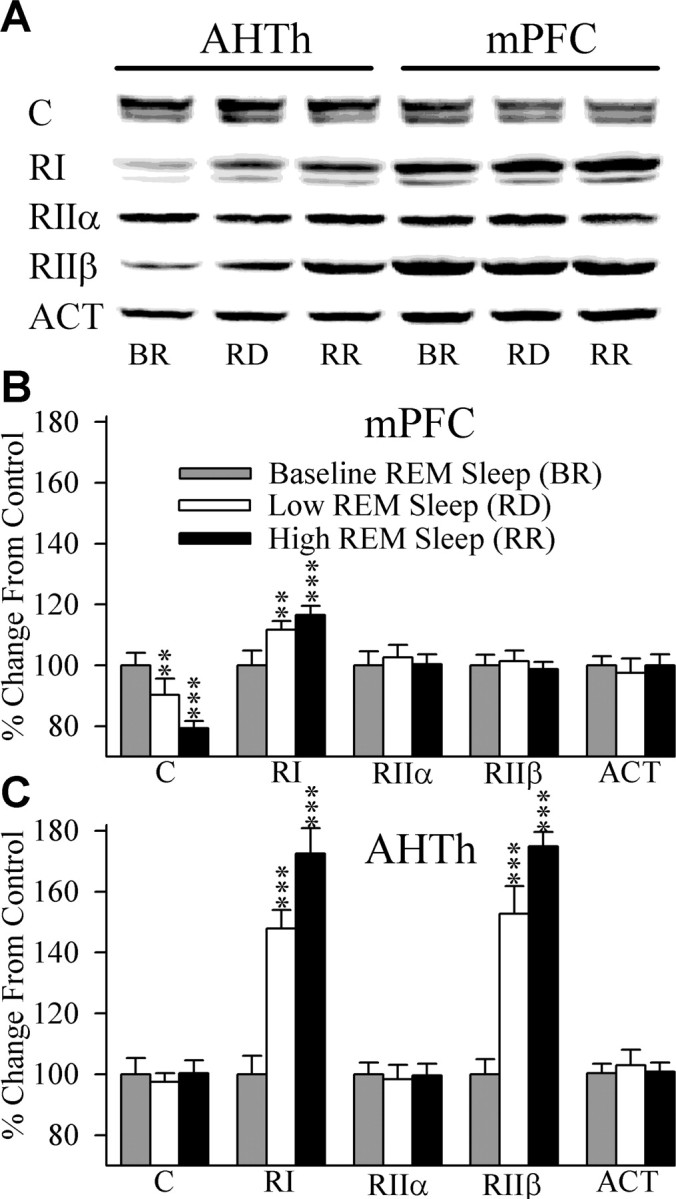
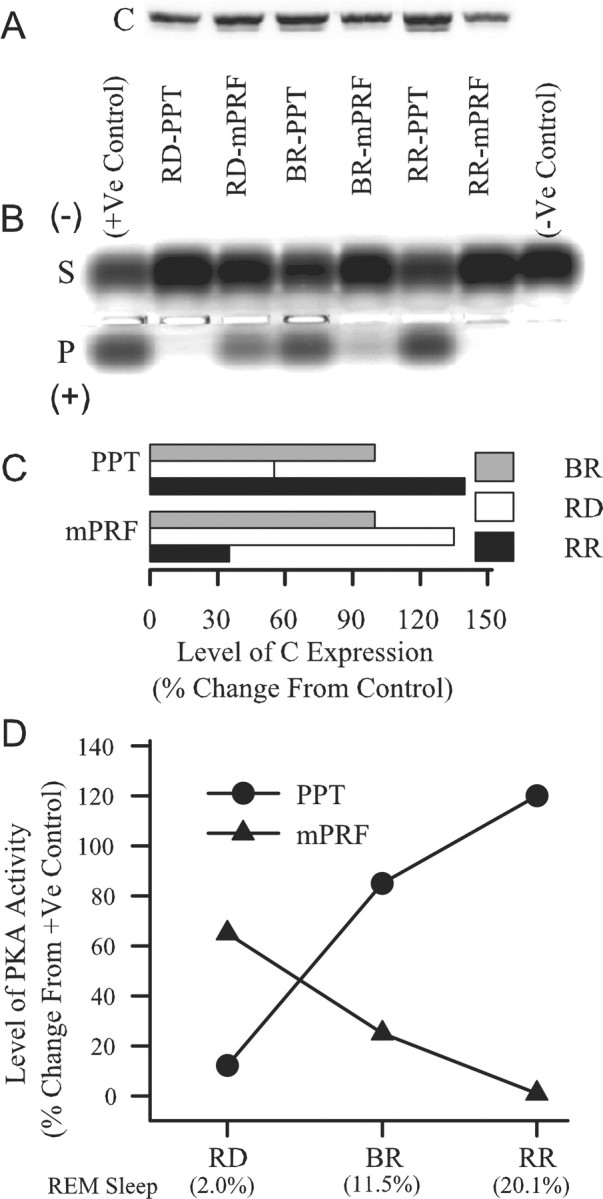
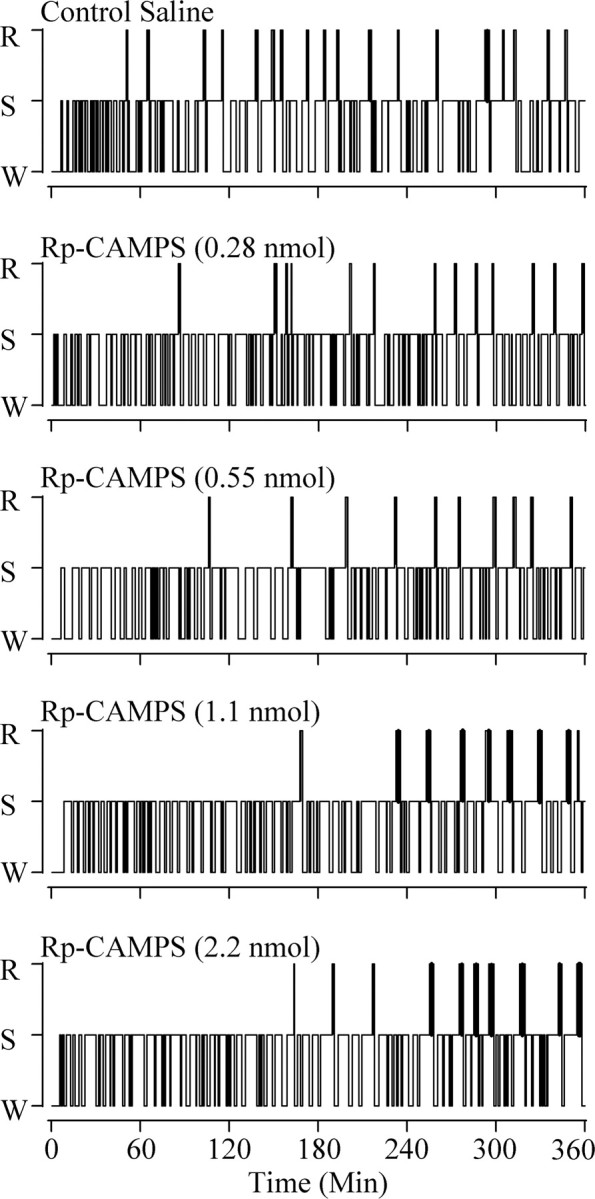
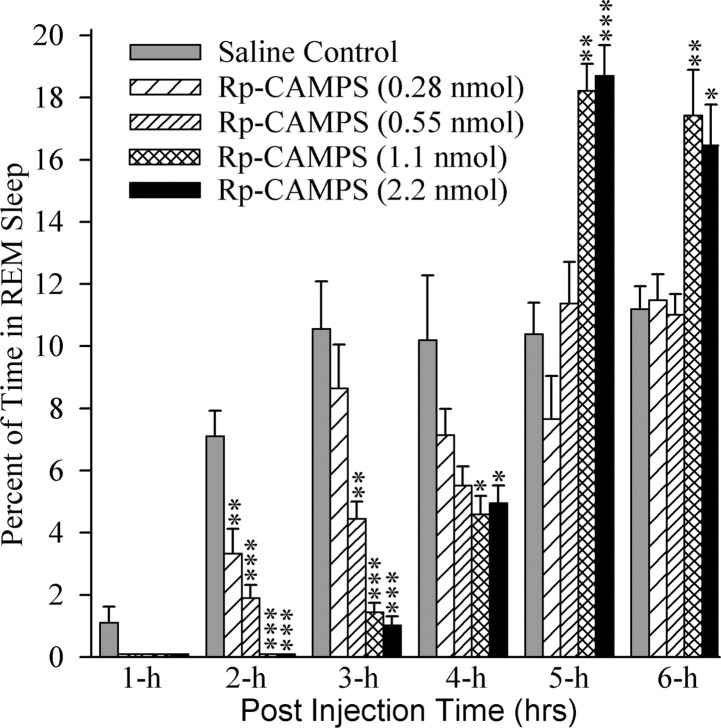
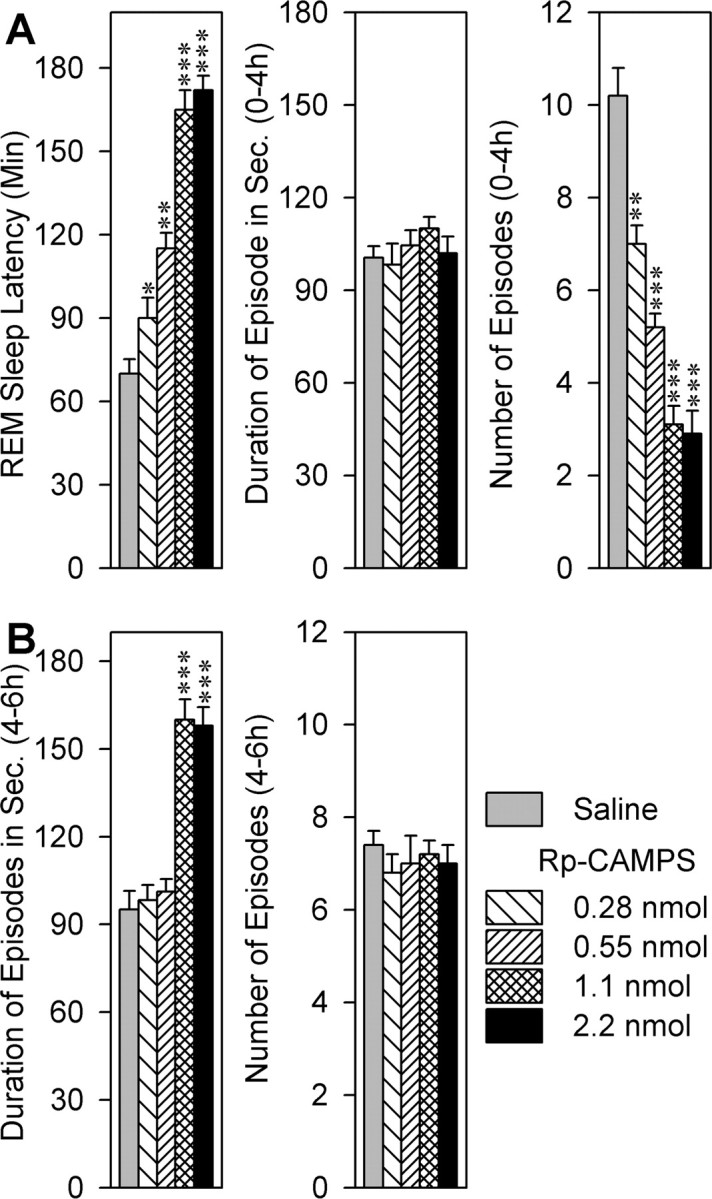
Cells in the pedunculopontine tegmentum (PPT) play a key role in the generation of rapid eye movement (REM) sleep, but its intracellular signaling mechanisms remain unknown. In the current studies, the role of PPT intracellular protein kinase A (PKA) in the regulation of REM sleep was evaluated by comparing PKA subunit [catalytic (PKA(C alpha)) and regulatory (PKA(RI), PKA(RII alpha), and PKA(RII beta)) types] expression and activity in the PPT at normal, high, and low REM sleep conditions. To compare anatomical specificity, REM sleep-dependent expressions of these PKA subunits were also measured in the medial pontine reticular formation (mPRF), medial prefrontal cortex (mPFC), and anterior hypothalamus (AHTh). The results of these PKA subunit expression and activity studies demonstrated that the expression of PKA(C alpha) and PKA activity in the PPT increased and decreased during high and low REM sleep, respectively. Conversely, PKA(C alpha) expression and PKA activity decreased with high REM sleep in the mPRF. Expression of PKA(C alpha) also decreased in the mPFC and remained unchanged in the AHTh with high REM sleep. These subunit expression and PKA activity data reveal a positive relationship between REM sleep and increased PKA activity in the PPT. To test this molecular evidence, localized activation of cAMP-dependent PKA activity was blocked using a pharmacological technique. The results of this pharmacological study demonstrated that the localized inhibition of cAMP-dependent PKA activation in the PPT dose-dependently suppressed REM sleep. Together, these results provide the first evidence that the activation of the PPT intracellular PKA system is involved in the generation of REM sleep.
Figures
References
-
- Ahnaou A, Basille M, Gonzalez B, Vaudry H, Hamon M, Adrien J, Bourgin P. Long-term enhancement of REM sleep by the pituitary adenylyl cyclase-activating polypeptide (PACAP) in the pontine reticular formation of rat. Eur J Neurosci. 1999;11:4051–4058. - PubMed
-
- Borrelli E, Montmayeur JP, Foulkes NS, Sassone-Corsi P. Signal transduction and gene control: the cAMP pathway. Crit Rev Oncog. 1992;3:321–338. - PubMed
-
- Bradford M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem. 1976;72:248–254. - PubMed
-
- Capece ML, Lydic R. cAMP and protein kinase A modulate cholinergic rapid eye movement sleep generation. Am J Physiol. 1997;273:R1430–R1440. - PubMed
-
- Cook SA, Welch SP, Lichtman AH, Martin BR. Evaluation of cAMP involvement in cannabinoid-induced antinociception. Life Sci. 1995;56:2049–2056. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources