

Prolongation of evoked and spontaneous synaptic currents at the neuromuscular junction after activity blockade is caused by the upregulation of fetal acetylcholine receptors
- PMID: 16943554
- PMCID: PMC6675328
- DOI: 10.1523/JNEUROSCI.2493-06.2006
Prolongation of evoked and spontaneous synaptic currents at the neuromuscular junction after activity blockade is caused by the upregulation of fetal acetylcholine receptors
Abstract
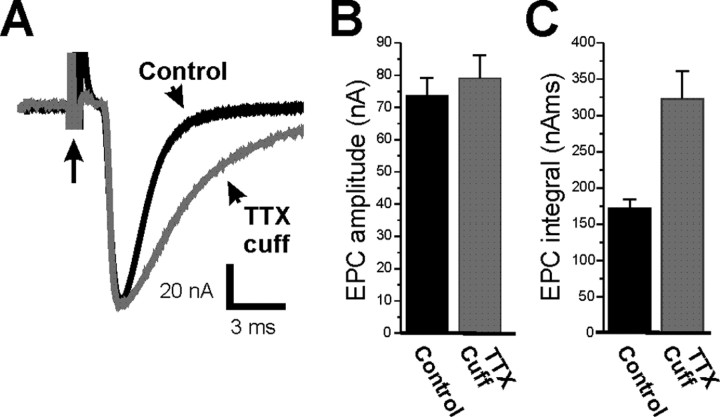
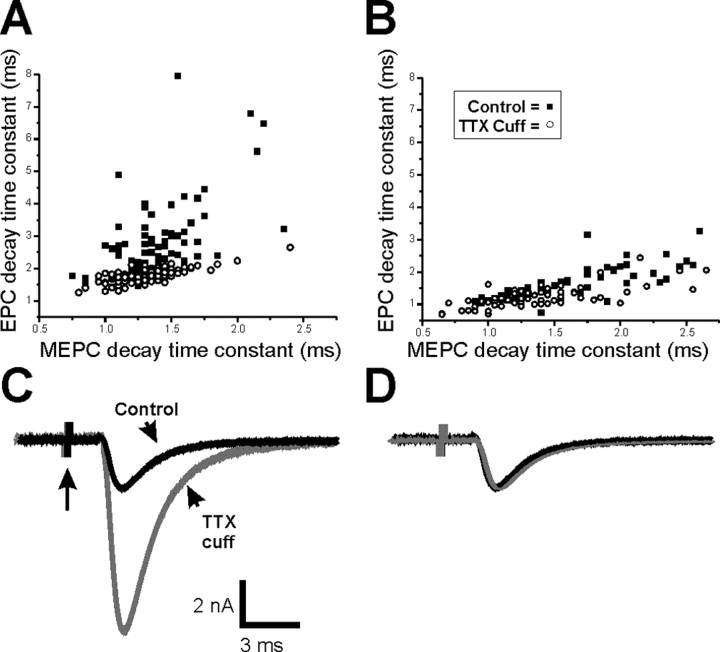
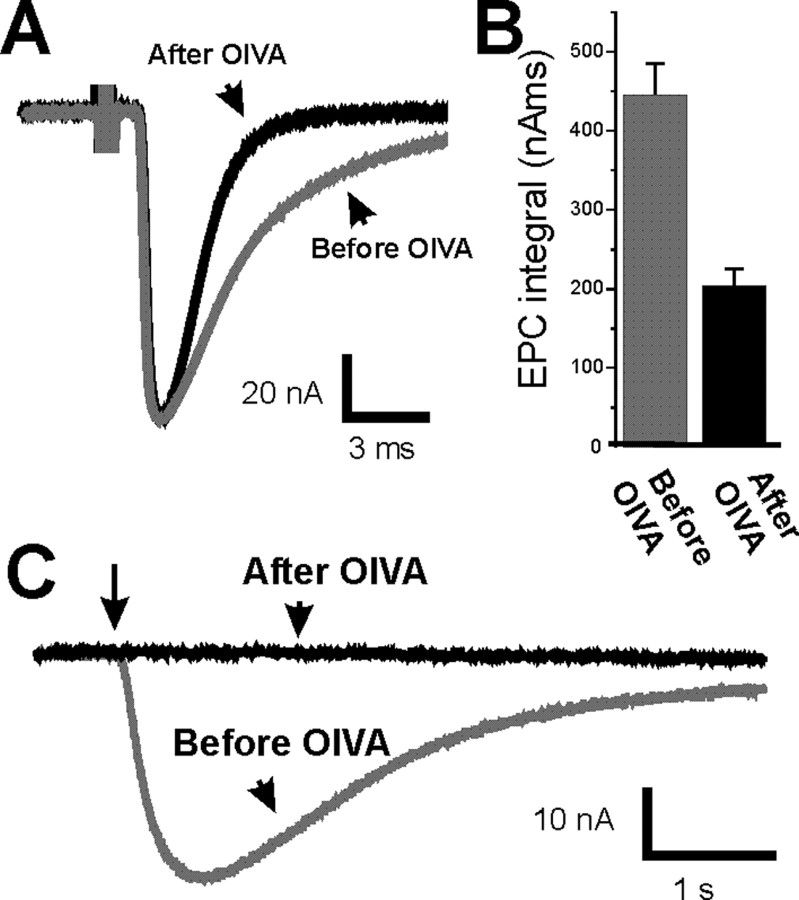
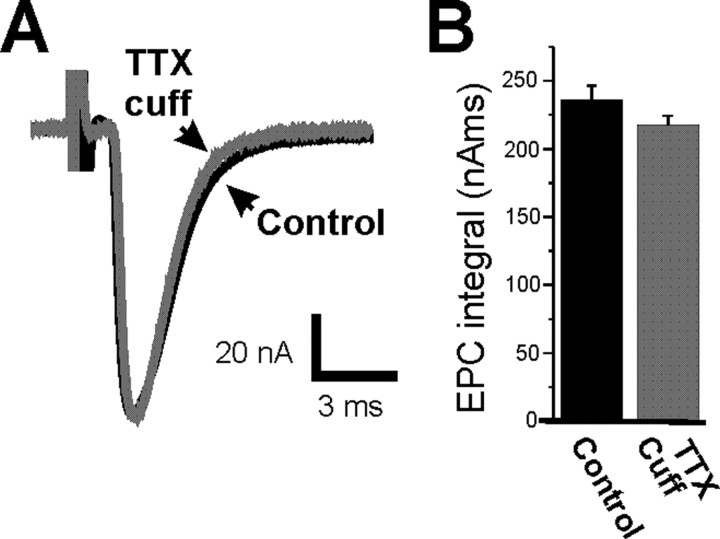
It has been shown previously in a number of systems that after an extended block of activity, synaptic strength is increased. We found that an extended block of synaptic activity at the mouse neuromuscular junction, using a tetrodotoxin cuff in vivo, increased synaptic strength by prolonging the evoked endplate current (EPC) decay. Prolongation of EPC decay was accompanied by only modest prolongation of spontaneous miniature EPC (MEPC) decay. Prolongation of EPC decay was reversed when quantal content was lowered by reducing extracellular calcium. These findings suggested that the cause of EPC prolongation was presynaptic in origin. However, when we acutely inhibited fetal-type acetylcholine receptors (AChRs) using a novel peptide toxin (alphaA-conotoxin OIVA[K15N]), prolongation of both EPC and MEPC decay were reversed. We also blocked synaptic activity in a mutant strain of mice in which persistent muscle activity prevents upregulation of fetal-type AChRs. In these mice, there was no prolongation of EPC decay. We conclude that upregulation of fetal-type AChRs after blocking synaptic activity causes modest prolongation of MEPC decay that is accompanied by much greater prolongation of EPC decay. This might occur if acetylcholine escapes from endplates and binds to extrajunctional fetal-type AChRs only during large, evoked EPCs. Our study is the first to demonstrate a functional role for upregulation of extrajunctional AChRs.
Figures
Similar articles
-
Synaptic depression in frog neuromuscular junction.J Neurophysiol. 1987 Jul;58(1):230-46. doi: 10.1152/jn.1987.58.1.230. J Neurophysiol. 1987. PMID: 2441003
-
Impaired synaptic vesicle release and immaturity of neuromuscular junctions in spinal muscular atrophy mice.J Neurosci. 2009 Jan 21;29(3):842-51. doi: 10.1523/JNEUROSCI.4434-08.2009. J Neurosci. 2009. PMID: 19158308 Free PMC article.
-
Metabolic stabilization of acetylcholine receptors at newly formed neuromuscular junctions in rat.Dev Biol. 1981 Jun;84(2):247-54. doi: 10.1016/0012-1606(81)90392-4. Dev Biol. 1981. PMID: 20737862
-
[Effect of blockade of axonal transport on endplate currents of muscle fibers in the frog].Neirofiziologiia. 1985;17(2):204-11. Neirofiziologiia. 1985. PMID: 2582281 Russian.
-
Effects of the aminoglycoside antibiotics, streptomycin and neomycin, on neuromuscular transmission. II. Postsynaptic considerations.J Pharmacol Exp Ther. 1983 Jun;225(3):496-502. J Pharmacol Exp Ther. 1983. PMID: 6306208
Cited by
-
Acetylcholinesterase deficiency contributes to neuromuscular junction dysfunction in type 1 diabetic neuropathy.Am J Physiol Endocrinol Metab. 2012 Aug 15;303(4):E551-61. doi: 10.1152/ajpendo.00622.2011. Epub 2012 Jun 26. Am J Physiol Endocrinol Metab. 2012. PMID: 22739110 Free PMC article.
-
Reduced survival of motor neuron (SMN) protein in motor neuronal progenitors functions cell autonomously to cause spinal muscular atrophy in model mice expressing the human centromeric (SMN2) gene.J Neurosci. 2010 Sep 8;30(36):12005-19. doi: 10.1523/JNEUROSCI.2208-10.2010. J Neurosci. 2010. PMID: 20826664 Free PMC article.
-
Activity-dependent regulation of the binomial parameters p and n at the mouse neuromuscular junction in vivo.J Neurophysiol. 2010 Nov;104(5):2352-8. doi: 10.1152/jn.00460.2010. Epub 2010 Aug 25. J Neurophysiol. 2010. PMID: 20739593 Free PMC article.
-
Distinct muscarinic acetylcholine receptor subtypes contribute to stability and growth, but not compensatory plasticity, of neuromuscular synapses.J Neurosci. 2009 Nov 25;29(47):14942-55. doi: 10.1523/JNEUROSCI.2276-09.2009. J Neurosci. 2009. PMID: 19940190 Free PMC article.
-
The Structure, Function, and Physiology of the Fetal and Adult Acetylcholine Receptor in Muscle.Front Mol Neurosci. 2020 Sep 8;13:581097. doi: 10.3389/fnmol.2020.581097. eCollection 2020. Front Mol Neurosci. 2020. PMID: 33013323 Free PMC article. Review.
References
-
- Argentieri TM, Aiken SP, Laxminarayan S, McArdle JJ. Characteristics of synaptic transmission in reinnervating rat skeletal muscle. Pflügers Arch. 1992;421:256–261. - PubMed
-
- Colomar A, Robitaille R. Glial modulation of synaptic transmission at the neuromuscular junction. Glia. 2004;47:284–289. - PubMed
-
- Davis GW. Homeostatic control of neural activity: from phenomenology to molecular design. Annu Rev Neurosci. 2006;29:307–323. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases