

Hair cell ribbon synapses
- PMID: 16944206
- PMCID: PMC4142044
- DOI: 10.1007/s00441-006-0276-3
Hair cell ribbon synapses
Abstract
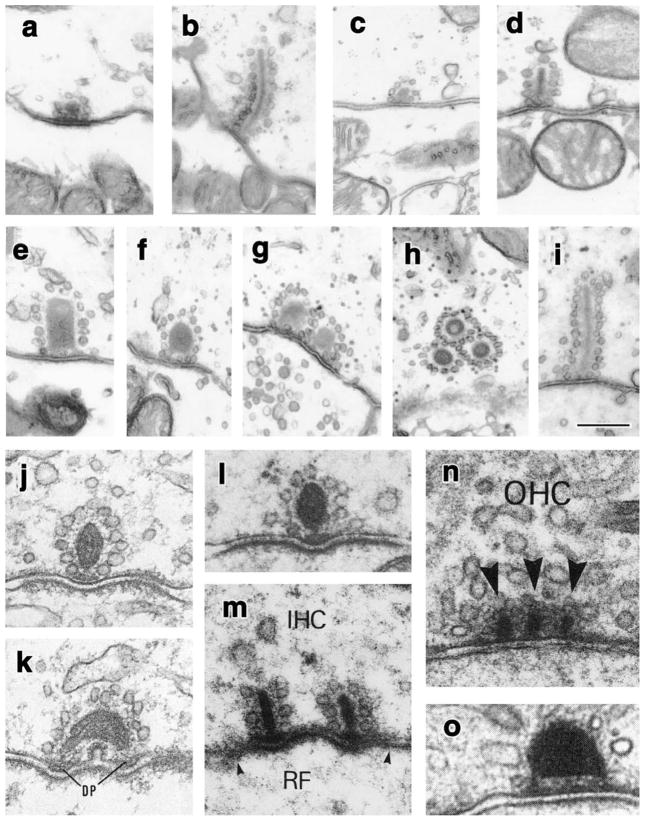
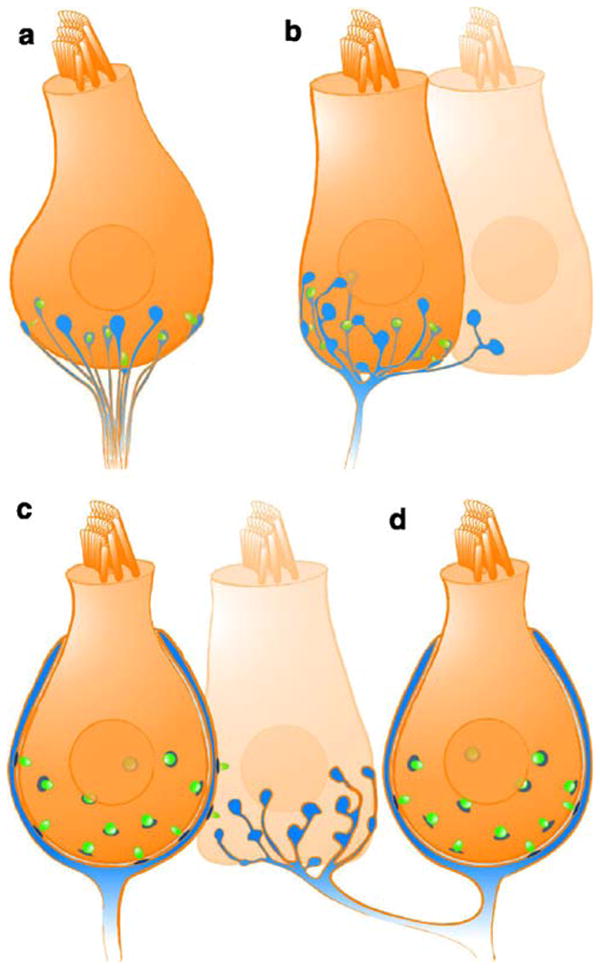
Hearing and balance rely on the faithful synaptic coding of mechanical input by the auditory and vestibular hair cells of the inner ear. Mechanical deflection of their stereocilia causes the opening of mechanosensitive channels, resulting in hair cell depolarization, which controls the release of glutamate at ribbon-type synapses. Hair cells have a compact shape with strong polarity. Mechanoelectrical transduction and active membrane turnover associated with stereociliar renewal dominate the apical compartment. Transmitter release occurs at several active zones along the basolateral membrane. The astonishing capability of the hair cell ribbon synapse for temporally precise and reliable sensory coding has been the subject of intense investigation over the past few years. This research has been facilitated by the excellent experimental accessibility of the hair cell. For the same reason, the hair cell serves as an important model for studying presynaptic Ca(2+) signaling and stimulus-secretion coupling. In addition to common principles, hair cell synapses differ in their anatomical and functional properties among species, among the auditory and vestibular organs, and among hair cell positions within the organ. Here, we briefly review synaptic morphology and connectivity and then focus on stimulus-secretion coupling at hair cell synapses.
Figures

References
-
- Art JJ, Crawford AC, Fettiplace R. Electrical resonance and membrane currents in turtle cochlear hair cells. Hear Res. 1986;22:31–36. - PubMed
-
- Augustine GJ. How does calcium trigger neurotransmitter release? Curr Opin Neurobiol. 2001;11:320–326. - PubMed
-
- Augustine GJ, Adler EM, Charlton MP. The calcium signal for transmitter secretion from presynaptic nerve terminals. Ann N Y Acad Sci. 1991;635:365–381. - PubMed
-
- Augustine GJ, Santamaria F, Tanaka K. Local calcium signaling in neurons. Neuron. 2003;40:331–346. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
