

Leishmania disease development depends on the presence of apoptotic promastigotes in the virulent inoculum
- PMID: 16945916
- PMCID: PMC1564231
- DOI: 10.1073/pnas.0600843103
Leishmania disease development depends on the presence of apoptotic promastigotes in the virulent inoculum
Erratum in
- Proc Natl Acad Sci U S A. 2006 Oct 31;103(44):16615
Abstract
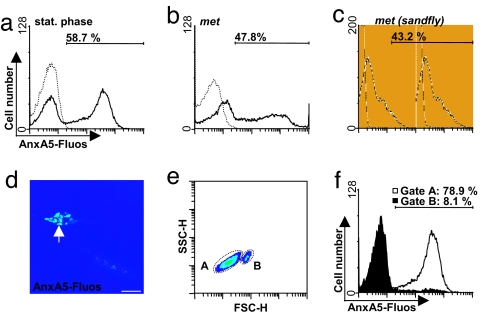
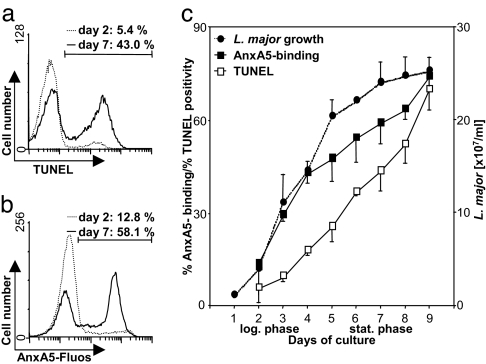
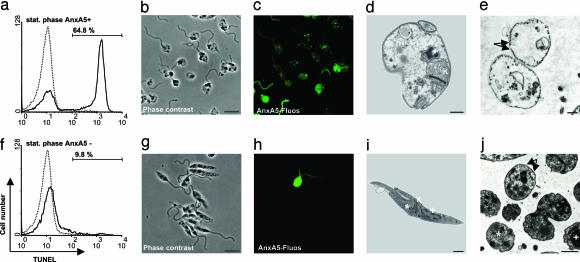
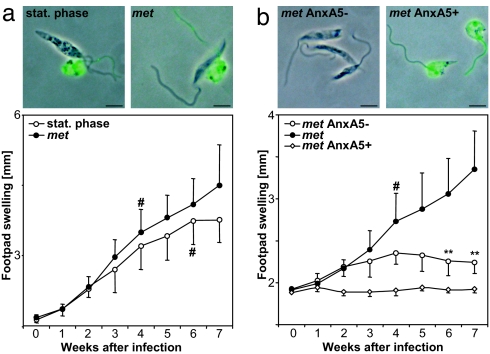
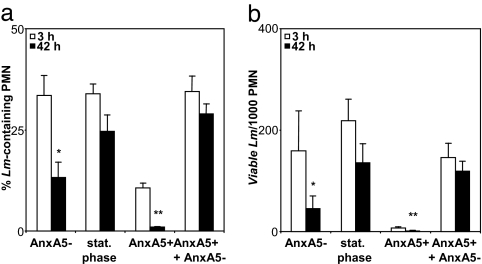
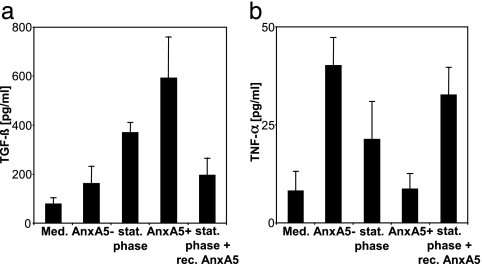
The obligate intracellular pathogen Leishmania major survives and multiplies in professional phagocytes. The evasion strategy to circumvent killing by host phagocytes and establish a productive infection is poorly understood. Here we report that the virulent inoculum of Leishmania promastigotes contains a high ratio of annexin A5-binding apoptotic parasites. This subpopulation of parasites is characterized by a round body shape, a swollen kinetoplast, nuclear condensation, and a lack of multiplication and represents dying or already dead parasites. After depleting the apoptotic parasites from a virulent population, Leishmania do not survive in phagocytes in vitro and lose their disease-inducing ability in vivo. TGF-beta induced by apoptotic parasites is likely to mediate the silencing of phagocytes and lead to survival of infectious Leishmania populations. The data demonstrate that apoptotic promastigotes, in an altruistic way, enable the intracellular survival of the viable parasites.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
