

Inhibition of a spliceosome turnover pathway suppresses splicing defects
- PMID: 16945917
- PMCID: PMC1564266
- DOI: 10.1073/pnas.0603188103
Inhibition of a spliceosome turnover pathway suppresses splicing defects
Abstract
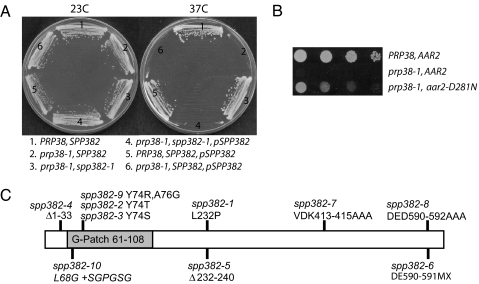
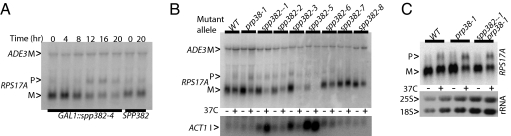
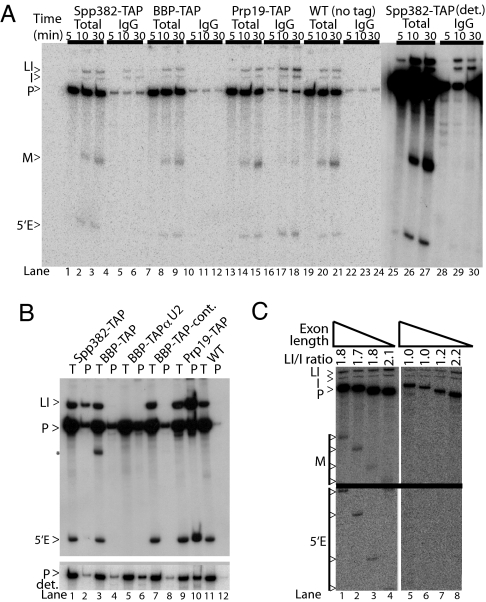
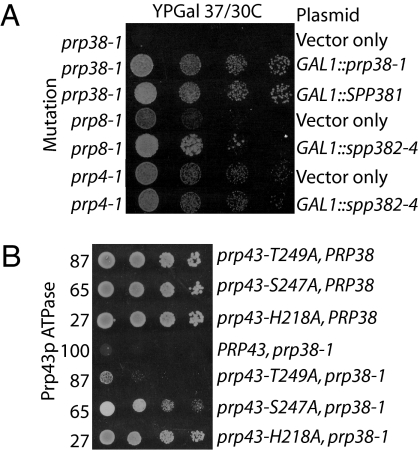
Defects in assembly are suggested to signal the dissociation of faulty splicing complexes. A yeast genetic screen was performed to identify components of the putative discard pathway. Weak mutant alleles of SPP382 (also called NTR1) were found to suppress defects in two proteins required for spliceosome activation, Prp38p and Prp8p. Spp382p is shown necessary for cellular splicing, with premRNA and, for some alleles, excised intron, accumulating after inactivation. Like spp382-1, a mutant allele of AAR2 was identified in this suppressor screen. Like Spp382p, Aar2p has a reported role in spliceosome recycling and is found with Spp382p in a complex recovered with a mutant version of the spliceosomal core protein Prp8p. Possible insight into to the spp382 suppressor phenotype is provided by the observation that defective splicing complexes lacking the 5' exon cleavage intermediate are recovered by a tandem affinity purification-tagged Spp382 derivative. Stringent proteomic and two-hybrid analyses show that Spp382p also interacts with Cwc23p, a DNA J-like protein present in the spliceosome and copurified with the Prp43p DExD/H-box ATPase. Spp382p binds Prp43p and Prp43p requires Spp382p for intron release from the spliceosome. Consistent with a related function in the removal of defective complexes, three prp43 mutants are also shown to suppress splicing defects, with efficiencies inversely proportionate to the measured ATPase activities. These and related genetic data support the existence of a Spp382p-dependent turnover pathway acting on defective spliceosomes.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
