

The echinoderm adhesome
- PMID: 16950242
- PMCID: PMC3565218
- DOI: 10.1016/j.ydbio.2006.07.044
The echinoderm adhesome
Abstract
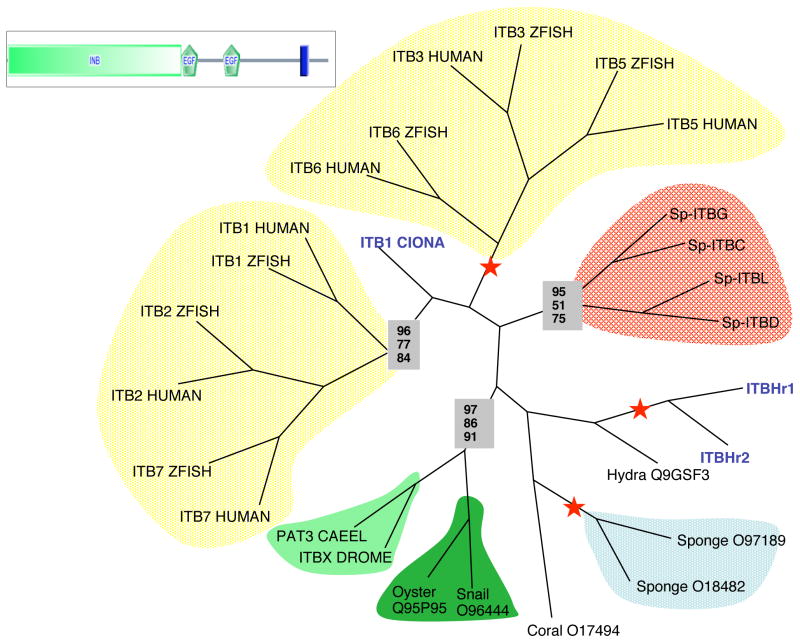
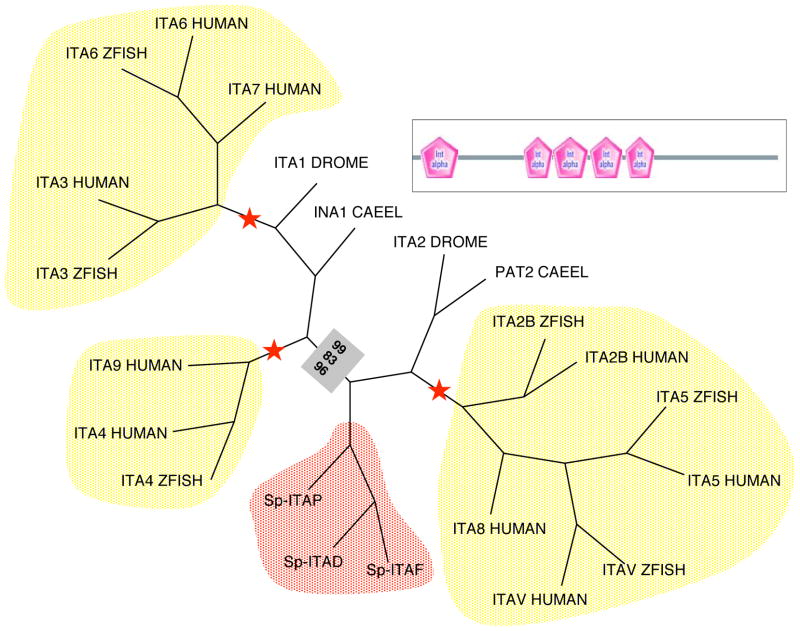
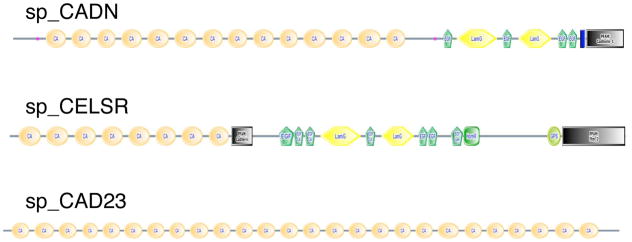
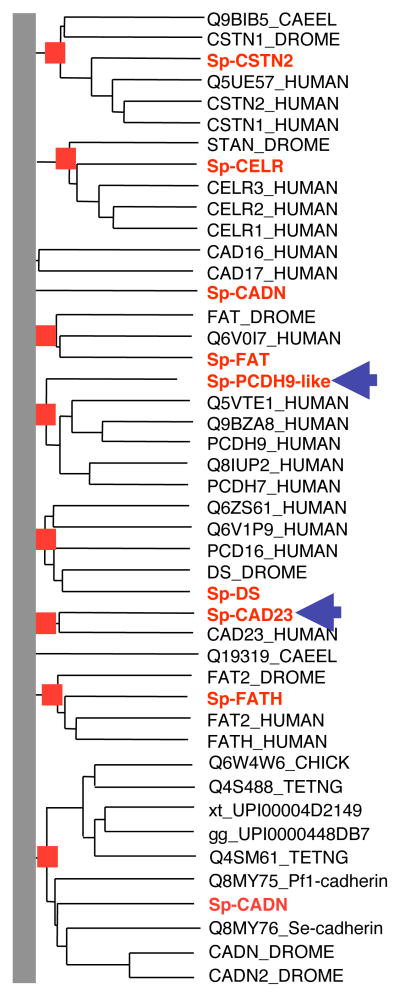
Although the development of sea urchin embryos has been studied extensively and clearly involves both cell adhesion and cell migration, rather little is known about the adhesion receptors and extracellular matrix molecules involved. The completion of the genome of Strongylocentrotus purpuratus allows a comprehensive survey of the complement of cell-cell and cell-matrix adhesion molecules in this organism. Furthermore, the phylogenetic position of echinoderms offers the opportunity to compare the complement of adhesion proteins between protostome and deuterostome invertebrates and between invertebrate and vertebrate deuterostomes. Many aspects of development and cell interactions differ among these different taxa and it is likely that analysis of the spectrum of adhesion receptors and extracellular matrix proteins can open up new insights into which molecules have evolved to suit particular developmental processes. In this paper, we report the results of an initial analysis along these lines. The echinoderm adhesome (complement of adhesion-related genes/proteins) is similar overall to that of other invertebrates although there are significant deuterostome-specific innovations and some interesting features previously thought to be chordate or vertebrate specific.
Figures
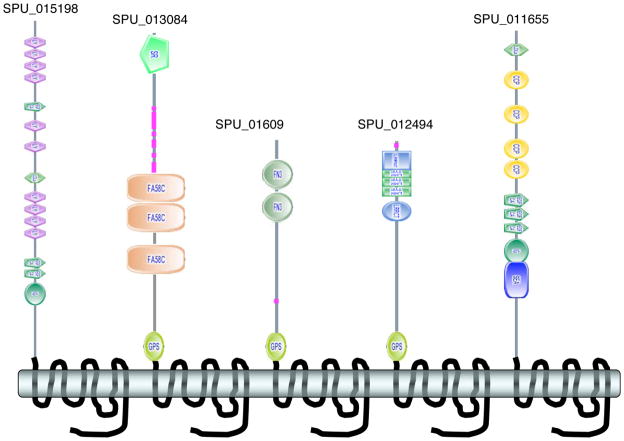
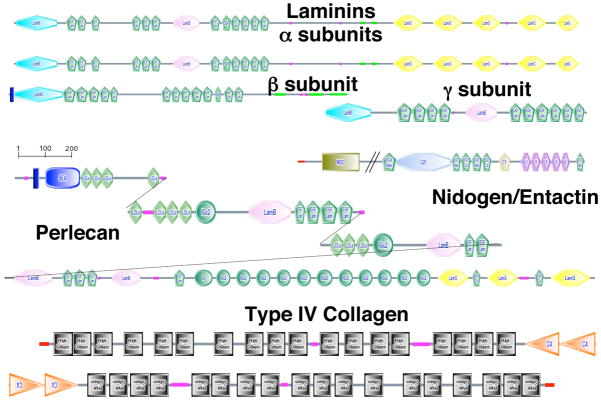
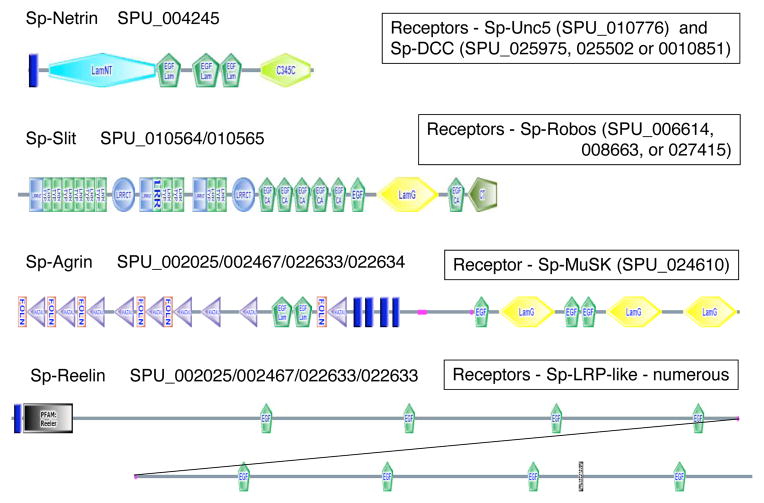
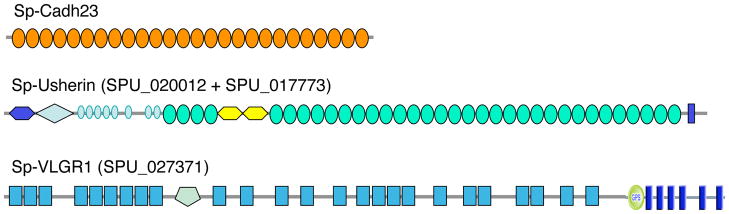
References
-
- Adato A, Lefevre G, Delprat B, Michel V, Michalski N, Chardenoux S, Weil D, El-Amraoui A, Petit C. Usherin, the defective protein in Usher syndrome type IIA, is likely to be a component of interstereocilia ankle links in the inner ear sensory cells. Hum Mol Genet. 2005a;14:3921–32. - PubMed
-
- Adato A, Michel V, Kikkawa Y, Reiners J, Alagramam KN, Weil D, Yonekawa H, Wolfrum U, El-Amraoui A, Petit C. Interactions in the network of Usher syndrome type 1 proteins. Hum Mol Genet. 2005b;14:347–56. - PubMed
-
- Behar O, Golden JA, Mashimo H, Schoen FJ, Fishman MC. Semaphorin III is needed for normal patterning and growth of nerves, bones and heart. Nature. 1996;383:525–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
