

Glutathione reductase and a mitochondrial thioredoxin play overlapping roles in maintaining iron-sulfur enzymes in fission yeast
- PMID: 16950927
- PMCID: PMC1694798
- DOI: 10.1128/EC.00244-06
Glutathione reductase and a mitochondrial thioredoxin play overlapping roles in maintaining iron-sulfur enzymes in fission yeast
Abstract
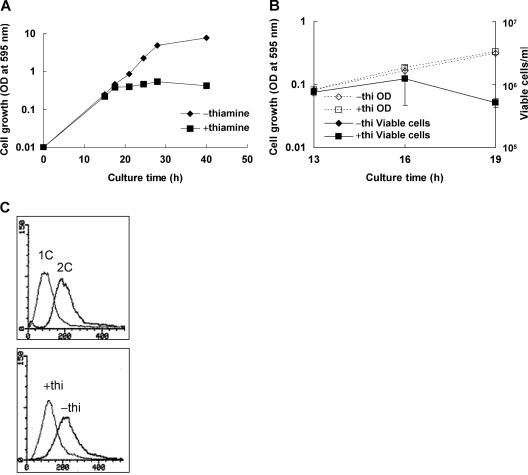
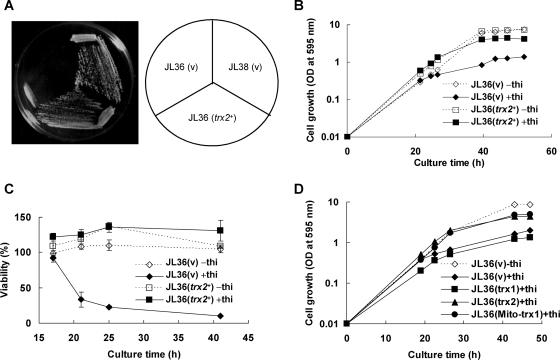
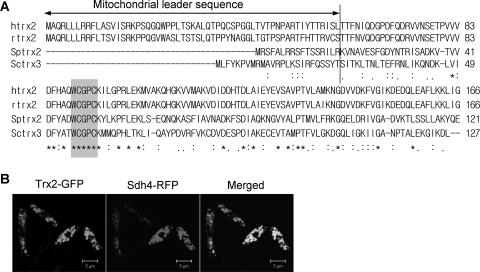
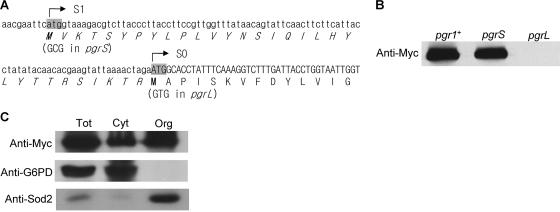
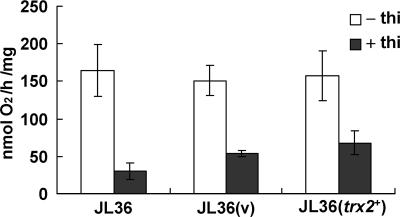
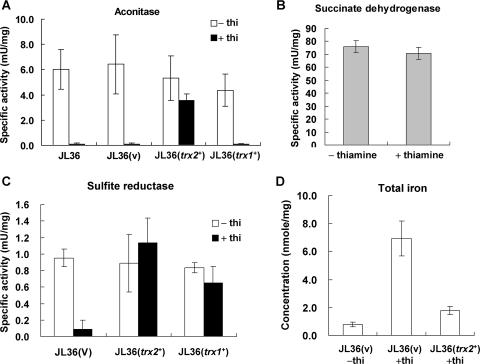
In the fission yeast Schizosaccharomyces pombe, the pgr1+ gene encoding glutathione (GSH) reductase (GR) is essentially required for cell survival. Depletion of GR caused proliferation arrest at the G1 phase of the cell cycle under aerobic conditions. Multicopy suppressors that restore growth were screened, and one effective suppressor was found to be the trx2+ gene, encoding a mitochondrial thioredoxin. This suggests that GR is critically required for some mitochondrial function(s). We found that GR resides in both cytosolic and organellar fractions of the cell. Depletion of GR lowered the respiration rate and the activity of oxidation-labile Fe-S enzymes such as mitochondrial aconitase and cytosolic sulfite reductase. Trx2 did not reverse the high ratio of oxidized glutathione to GSH or the low respiration rate observed in GR-depleted cells. However, it brought the activity of oxidation-labile Fe-S enzymes to a normal level, suggesting that the maintenance of Fe-S enzymes is a critical factor in the survival of S. pombe. The activity of succinate dehydrogenase, an oxidation-insensitive Fe-S enzyme, however, was not affected by GR depletion, suggesting that GR is not required for the biogenesis of the Fe-S cluster. The total iron content was greatly increased by GR depletion and was brought to a nearly normal level by Trx2. These results indicate that the essentiality of GR in the aerobic growth of S. pombe is derived from its role in maintaining oxidation-labile Fe-S enzymes and iron homeostasis.
Figures
References
-
- Alfa, C., P. Fantes, J. Hyams, M. Mcleod, and E. Warbrick. 1993. Experiments with fission yeast: a laboratory course manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
-
- Beer, S. M., E. R. Taylor, S. E. Brown, C. C. Dahm, N. J. Costa, M. J. Runswick, and M. P. Murphy. 2004. Glutaredoxin 2 catalyzes the reversible oxidation and glutathionylation of mitochondrial membrane thiol proteins: implications for mitochondrial redox regulation and antioxidant defense. J. Biol. Chem. 279:47939-47951. - PubMed
-
- Beinert, H., R. H. Holm, and E. Münck. 1997. Iron-sulfur clusters: nature's modular, multipurpose structures. Science 277:653-659. - PubMed
-
- Beinert, H., and P. J. Kiley. 1999. Fe-S proteins in sensing and regulatory functions. Curr. Opin. Chem. Biol. 3:152-157. - PubMed
-
- Cadenas, E., and K. J. A. Davies. 2000. Mitochondrial free radical generation, oxidative stress, and aging. Free Radic. Biol. Med. 29:222-230. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
