

The Pseudomonas aeruginosa universal stress protein PA4352 is essential for surviving anaerobic energy stress
- PMID: 16952944
- PMCID: PMC1595484
- DOI: 10.1128/JB.00308-06
The Pseudomonas aeruginosa universal stress protein PA4352 is essential for surviving anaerobic energy stress
Abstract
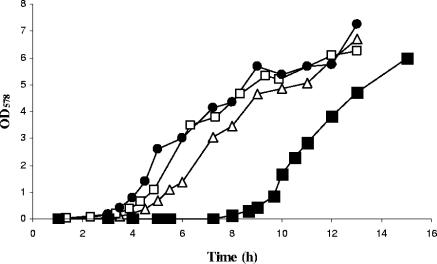
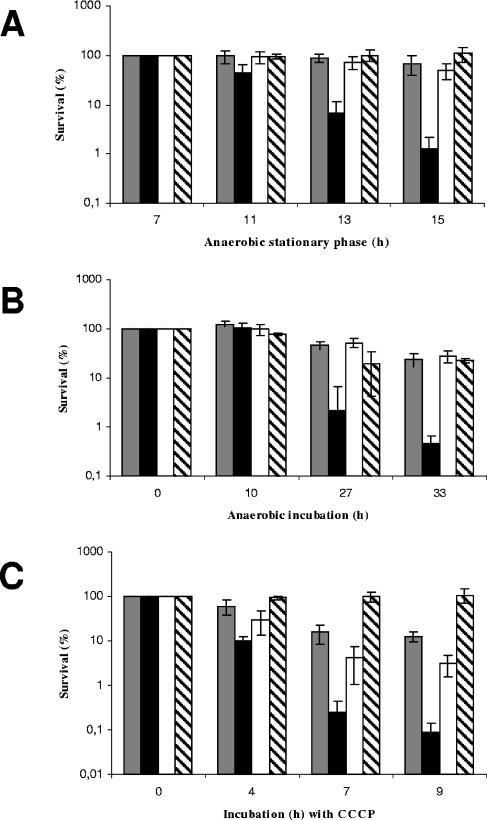
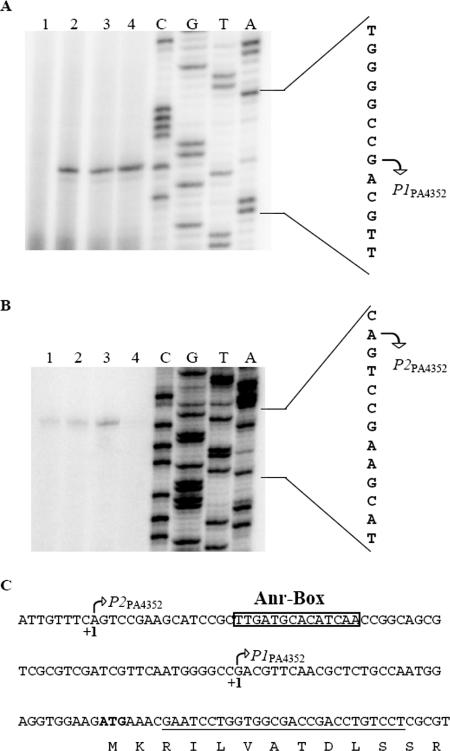
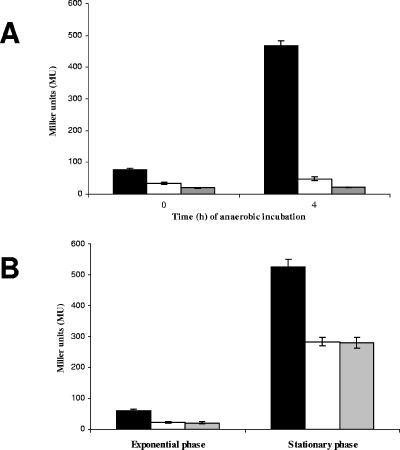
During infection of the cystic fibrosis (CF) lung, Pseudomonas aeruginosa microcolonies are embedded in the anaerobic CF mucus. This anaerobic environment seems to contribute to the formation of more robust P. aeruginosa biofilms and to an increased antibiotic tolerance and therefore promotes persistent infection. This study characterizes the P. aeruginosa protein PA4352, which is important for survival under anaerobic energy stress conditions. PA4352 belongs to the universal stress protein (Usp) superfamily and harbors two Usp domains in tandem. In Escherichia coli, Usp-type stress proteins are involved in survival during aerobic growth arrest and under various other stresses. A P. aeruginosa PA4352 knockout mutant was tested for survival under several stress conditions. We found a decrease in viability of this mutant compared to the P. aeruginosa wild type during anaerobic energy starvation caused by the missing electron acceptors oxygen and nitrate. Consistent with this phenotype under anaerobic conditions, the PA4352 knockout mutant was also highly sensitive to carbonyl cyanide m-chlorophenylhydrazone, the chemical uncoupler of the electron transport chain. Primer extension experiments identified two promoters upstream of the PA4352 gene. One promoter is activated in response to oxygen limitation by the oxygen-sensing regulatory protein Anr. The center of a putative Anr binding site was identified 41.5 bp upstream of the transcriptional start site. The second promoter is active only in the stationary phase, however, independently of RpoS, RelA, or quorum sensing. This is the second P. aeruginosa Usp-type stress protein that we have identified as important for survival under anaerobic conditions, which resembles the environment during persistent infection.
Figures

References
-
- Aiba, H., S. Adhya, and B. de Crombrugghe. 1981. Evidence for two gal promoters in intact Escherichia coli. J. Biol. Chem. 256:11905-11910. - PubMed
-
- Arai, H., T. Kodama, and Y. Igarashi. 1997. Cascade regulation of the two CRP/FNR-related transcriptional regulators (ANR and DNR) and the denitrification enzymes in Pseudomonas aeruginosa. Mol. Microbiol. 25:1141-1148. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl. 1995. Current protocols in molecular biology. John Wiley & Sons, New York, N.Y.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
