

Hypoxanthine incorporation is nonmutagenic in Escherichia coli
- PMID: 16952947
- PMCID: PMC1595496
- DOI: 10.1128/JB.00447-06
Hypoxanthine incorporation is nonmutagenic in Escherichia coli
Abstract
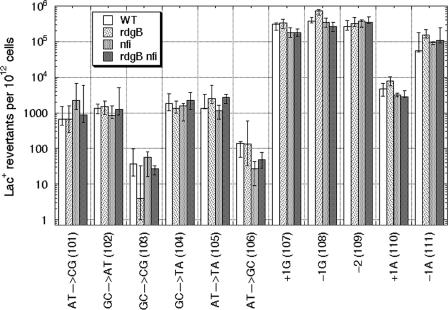
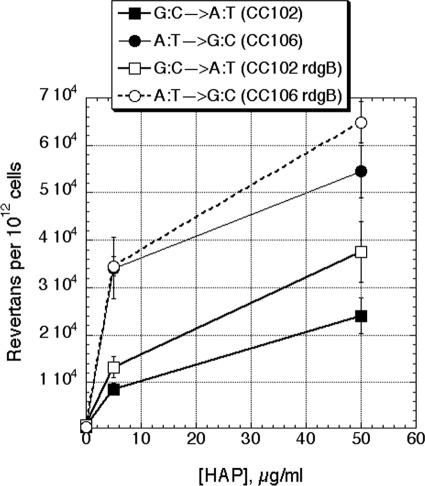
Endonuclease V, encoded by the nfi gene, initiates removal of the base analogs hypoxanthine and xanthine from DNA, acting to prevent mutagenesis from purine base deamination within the DNA. On the other hand, the RdgB nucleotide hydrolase in Escherichia coli is proposed to prevent hypoxanthine and xanthine incorporation into DNA by intercepting the noncanonical DNA precursors dITP and dXTP. Because many base analogs are mutagenic when incorporated into DNA, it is intuitive to think of RdgB as acting to prevent similar mutagenesis from deaminated purines in the DNA precursor pools. To test this idea, we used a set of Claire Cupples' strains to detect changes in spontaneous mutagenesis spectra, as well as in nitrous acid-induced mutagenesis spectra, in wild-type cells and in rdgB single, nfi single, and rdgB nfi double mutants. We found neither a significant increase in spontaneous mutagenesis in rdgB and nfi single mutants or the double mutant nor any changes in nitrous acid-induced mutagenesis for rdgB mutant strains. We conclude that incorporation of deaminated purines into DNA is nonmutagenic.
Figures






References
-
- Abdul-Masih, M. T., and M. J. Bessman. 1986. Biochemical studies on the mutagen, 6-N-hydroxylaminopurine. Synthesis of the deoxynucleoside triphosphate and its incorporation into DNA in vitro. J. Biol. Chem. 261:2020-2026. - PubMed
-
- Bertani, L. E., A. Haggmark, and P. Reichard. 1963. Enzymatic synthesis of deoxyribonucleotides, II. Formation and interconversion of deoxyuridine phosphates. J. Biol. Chem. 238:3407-3413. - PubMed
-
- Birnboim, H. C. 1983. A rapid alkaline extraction method for the isolation of plasmid DNA. Methods Enzymol. 100:243-255. - PubMed
-
- Bradshaw, J. S., and A. Kuzminov. 2003. RdgB acts to avoid chromosome fragmentation in Escherichia coli. Mol. Microbiol. 48:1711-1725. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
