

Control of enterotoxin gene expression in Bacillus cereus F4430/73 involves the redox-sensitive ResDE signal transduction system
- PMID: 16952956
- PMCID: PMC1595479
- DOI: 10.1128/JB.00702-06
Control of enterotoxin gene expression in Bacillus cereus F4430/73 involves the redox-sensitive ResDE signal transduction system
Erratum in
- J Bacteriol. 2007 Mar;189(6):2581
Abstract
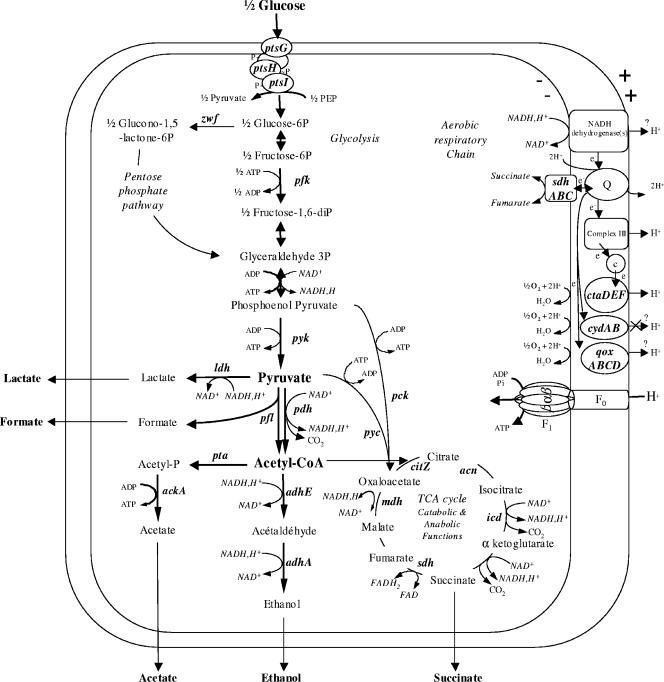
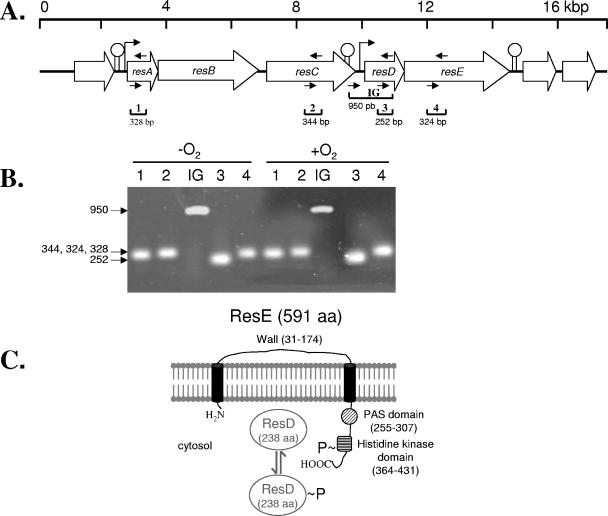
In contrast to Bacillus subtilis, the role of the two-component regulatory system ResDE has not yet been investigated in the facultative anaerobe Bacillus cereus. We examined the role of ResDE in the food-borne pathogen B. cereus F4430/73 by constructing resDE and resE mutants. Growth performances, glucose metabolism, and expression of hemolysin BL (Hbl) and nonhemolytic enterotoxin (Nhe) were analyzed in the three strains under distinct oxygenation and extracellular oxidoreduction potential (ORP) conditions. We show that growth and glucose metabolism were only moderately perturbed in both resDE and resE mutants under aerobiosis, microaerobiosis, and anaerobiosis generated under N(2) atmosphere (initial ORP = +45 mV). The major effects of resDE and resE mutations were observed under low-ORP anaerobic conditions generated under hydrogen atmosphere (iORP = -148 mV). These conditions normally favor enterotoxin production in the wild type. The resE mutation was more deleterious to the cells than the resDE mutation, causing growth limitation and strong deregulation of key catabolic genes. More importantly, the resE mutation abolished the production of enterotoxins under all of the conditions examined. The resDE mutation only decreased enterotoxin expression under anaerobiosis, with a more pronounced effect under low-ORP conditions. Thus, the ResDE system was found to exert major control on both fermentative growth and enterotoxin expression, and it is concluded that the ResDE system of B. cereus should be considered an anaerobic redox regulator. The data presented also provide evidence that the ResDE-dependent regulation of enterotoxins might function at least partially independently of the pleiotropic virulence gene regulator PlcR.
Figures
References
-
- Agaisse, H., M. Gominet, O. A. Okstad, A. B. Kolsto, and D. Lereclus. 1999. PlcR is a pleiotropic regulator of extracellular virulence factor gene expression in Bacillus thuringiensis. Mol. Microbiol. 32:1043-1053. - PubMed
-
- Andersson, A., P. E. Granum, and U. Ronner. 1998. The adhesion of Bacillus cereus spores to epithelial cells might be an additional virulence mechanism. Int. J. Food Microbiol. 39:93-99. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
