

Efficient loading of dendritic cells following cryo and radiofrequency ablation in combination with immune modulation induces anti-tumour immunity
- PMID: 16953240
- PMCID: PMC2360548
- DOI: 10.1038/sj.bjc.6603341
Efficient loading of dendritic cells following cryo and radiofrequency ablation in combination with immune modulation induces anti-tumour immunity
Abstract
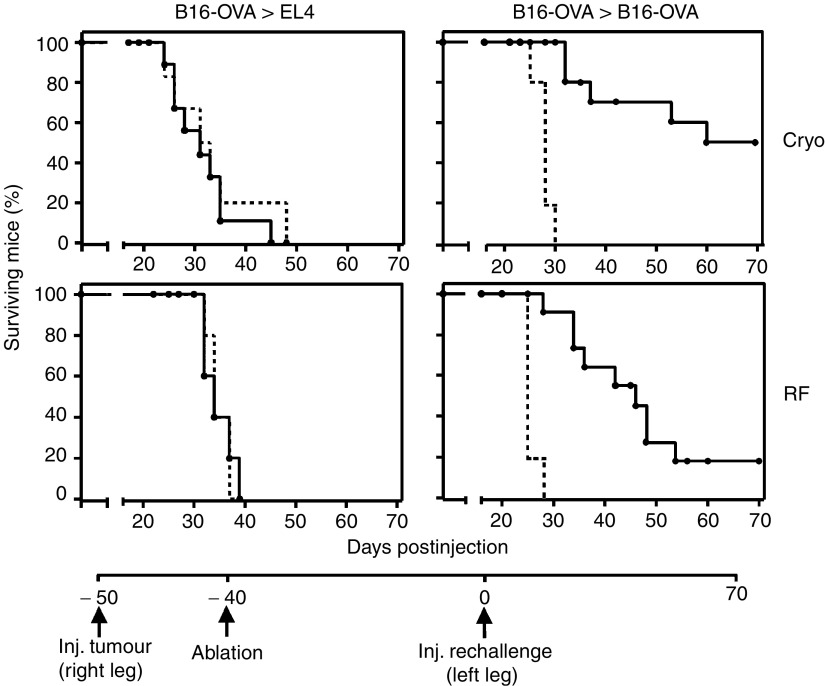
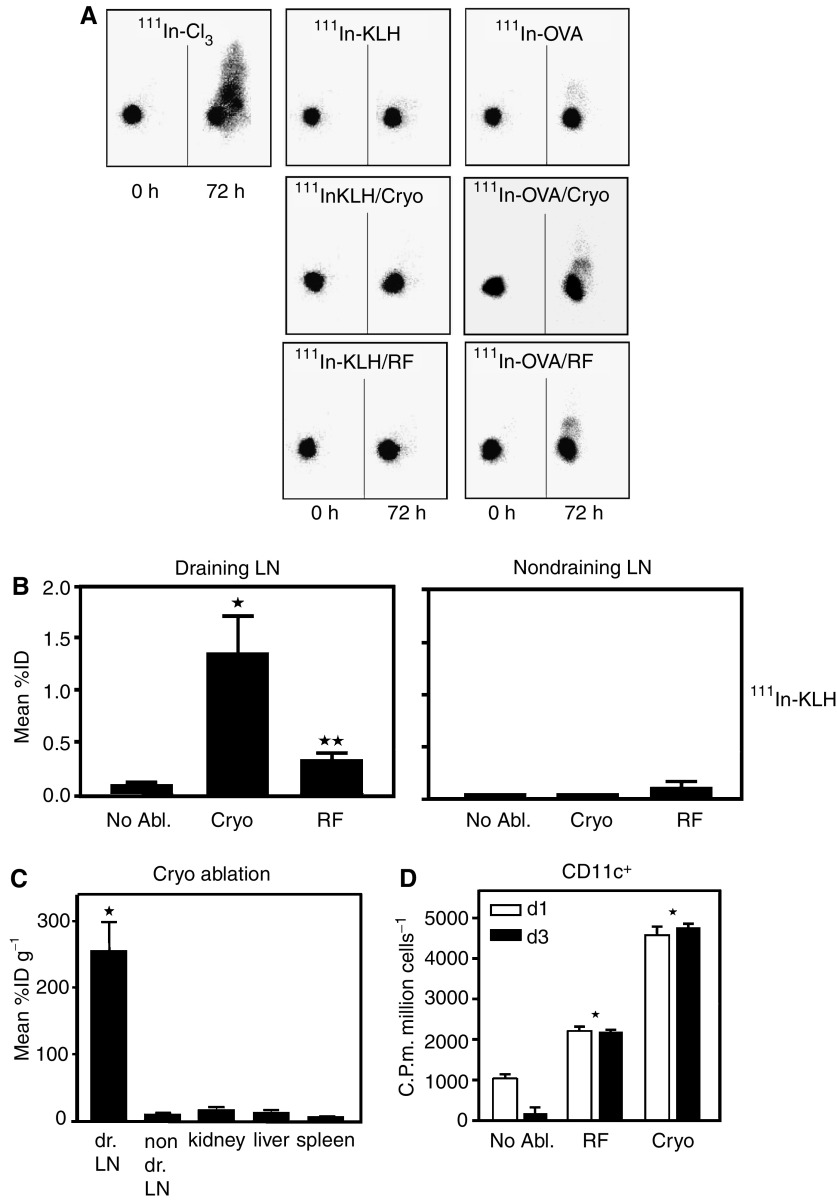
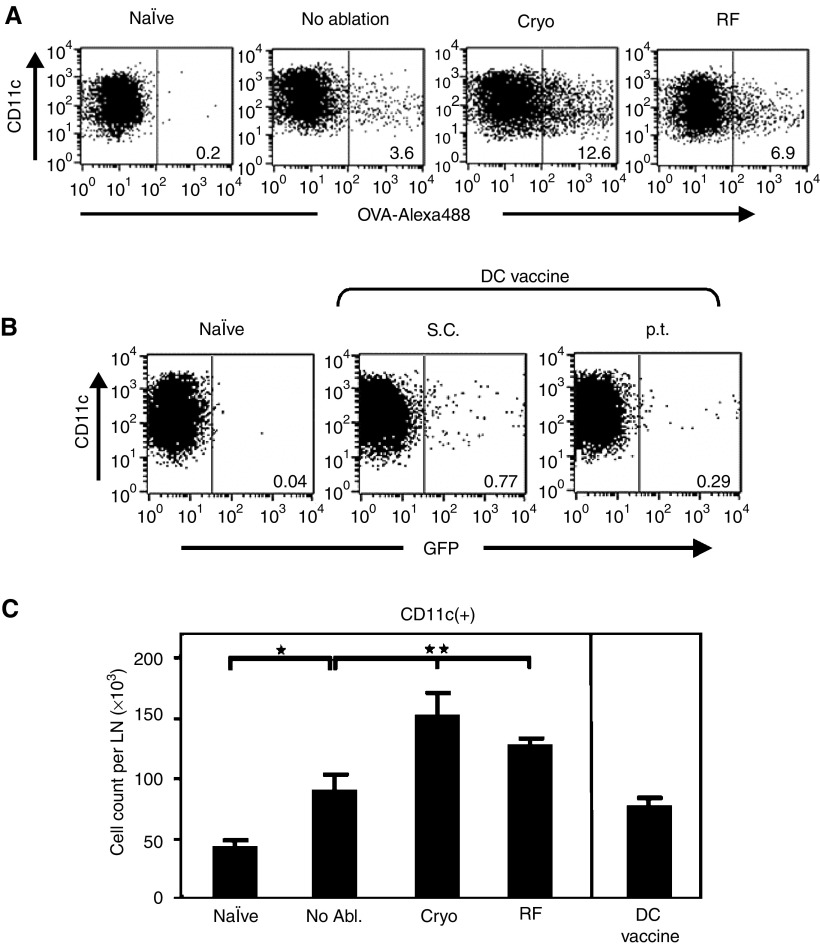
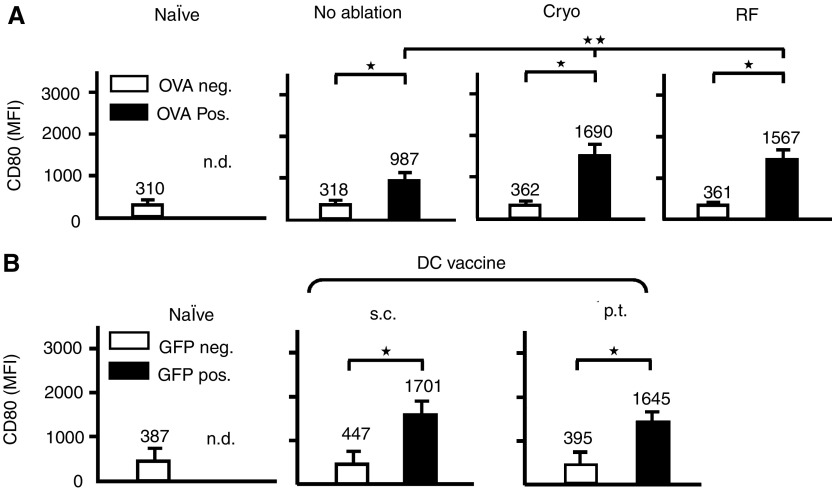
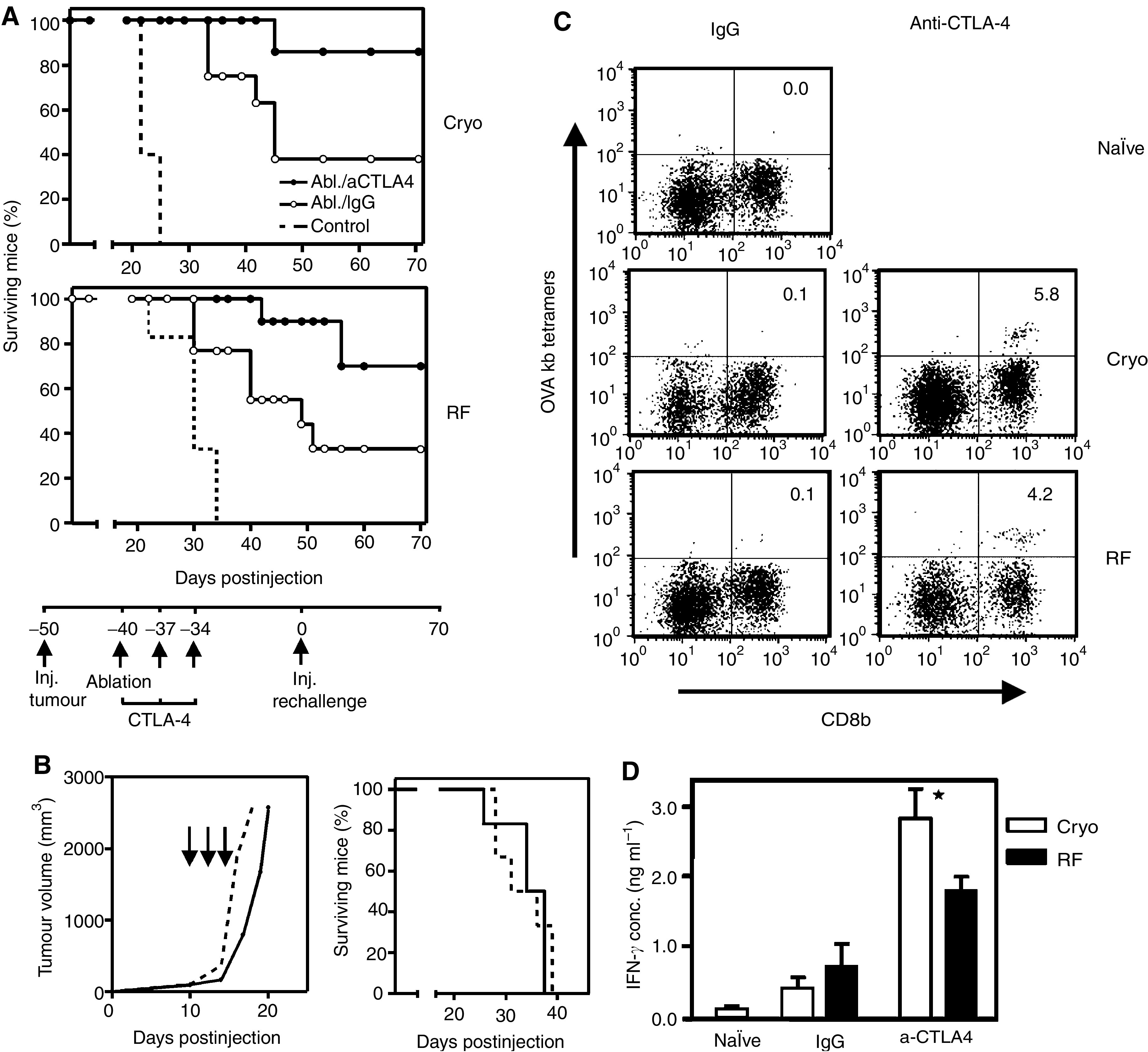
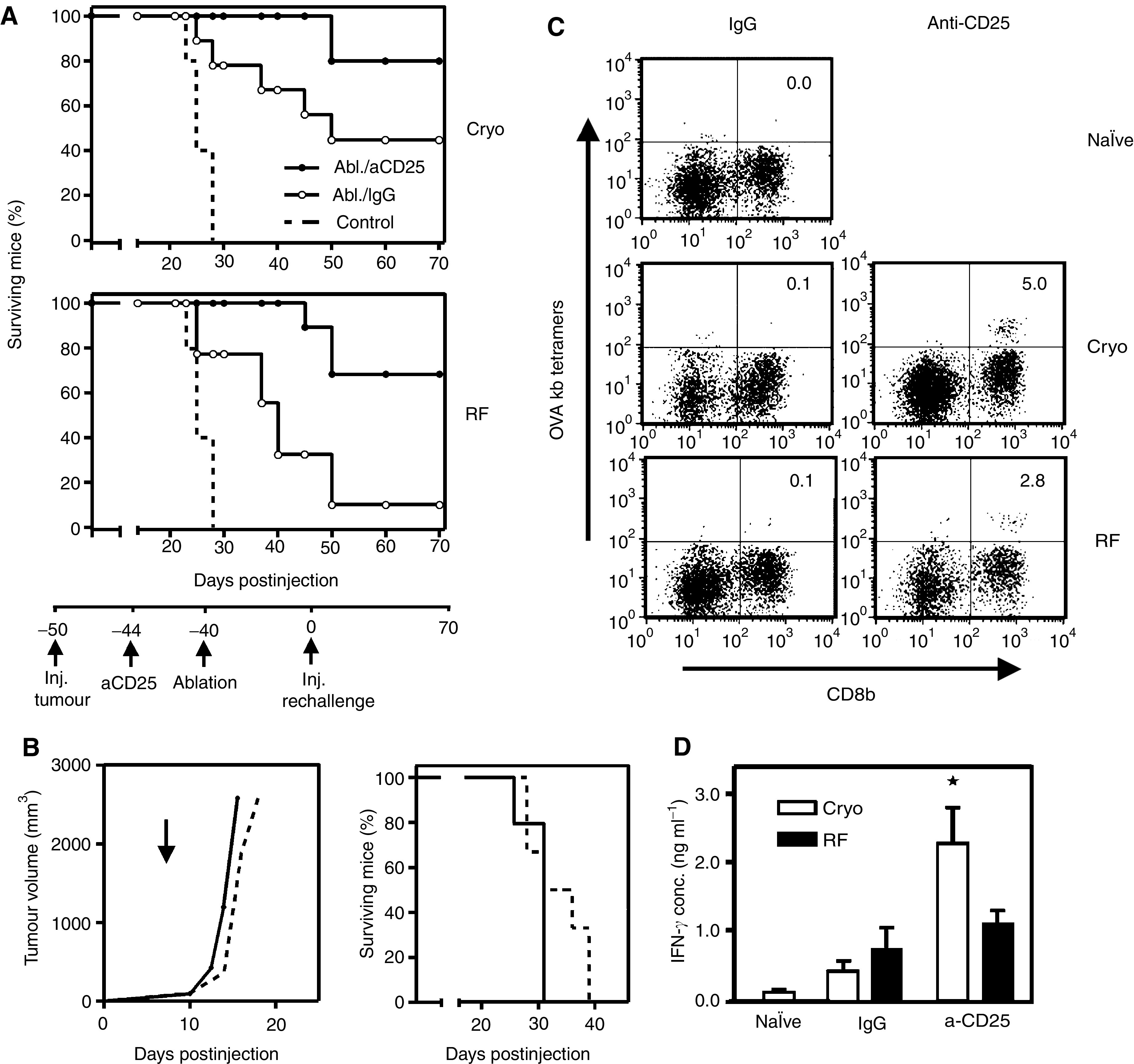
Dendritic cells (DC) are professional antigen-presenting cells that play a pivotal role in the induction of immunity. Ex vivo-generated, tumour antigen-loaded mature DC are currently exploited as cancer vaccines in clinical studies. However, antigen loading and maturation of DC directly in vivo would greatly facilitate the application of DC-based vaccines. We formerly showed in murine models that radiofrequency-mediated tumour destruction can provide an antigen source for the in vivo induction of anti-tumour immunity, and we explored the role of DC herein. In this paper we evaluate radiofrequency and cryo ablation for their ability to provide an antigen source for DC and compare this with an ex vivo-loaded DC vaccine. The data obtained with model antigens demonstrate that upon tumour destruction by radiofrequency ablation, up to 7% of the total draining lymph node (LN) DC contained antigen, whereas only few DC from the conventional vaccine reached the LN. Interestingly, following cryo ablation the amount of antigen-loaded DC is almost doubled. Analysis of surface markers revealed that both destruction methods were able to induce DC maturation. Finally, we show that in situ tumour ablation can be efficiently combined with immune modulation by anti-CTLA-4 antibodies or regulatory T-cell depletion. These combination treatments protected mice from the outgrowth of tumour challenges, and led to in vivo enhancement of tumour-specific T-cell numbers, which produced more IFN-gamma upon activation. Therefore, in situ tumour destruction in combination with immune modulation creates a unique, 'in situ DC-vaccine' that is readily applicable in the clinic without prior knowledge of tumour antigens.
Figures
References
-
- Adams S, O'Neill DW, Bhardwaj N (2005) Recent advances in dendritic cell biology. J Clin Immunol 25: 87–98 - PubMed
-
- Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka K (2000) Immunobiology of dendritic cells. Annu Rev Immunol 18: 767–811 - PubMed
-
- Banchereau J, Palucka AK (2005) Dendritic cells as therapeutic vaccines against cancer. Nat Rev Immunol 5: 296–306 - PubMed
-
- Chen L, Ashe S, Brady WA, Hellstrom I, Hellstrom KE, Ledbetter JA, McGowan P, Linsley PS (1992) Costimulation of antitumor immunity by the B7 counterreceptor for the T lymphocyte molecules CD28 and CTLA-4. Cell 71: 1093–1102 - PubMed
-
- Coulie PG, van der Bruggen P (2003) T-cell responses of vaccinated cancer patients. Curr Opin Immunol 15: 131–137 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
