

Structure of the DPS-like protein from Sulfolobus solfataricus reveals a bacterioferritin-like dimetal binding site within a DPS-like dodecameric assembly
- PMID: 16953567
- PMCID: PMC1815386
- DOI: 10.1021/bi060782u
Structure of the DPS-like protein from Sulfolobus solfataricus reveals a bacterioferritin-like dimetal binding site within a DPS-like dodecameric assembly
Abstract
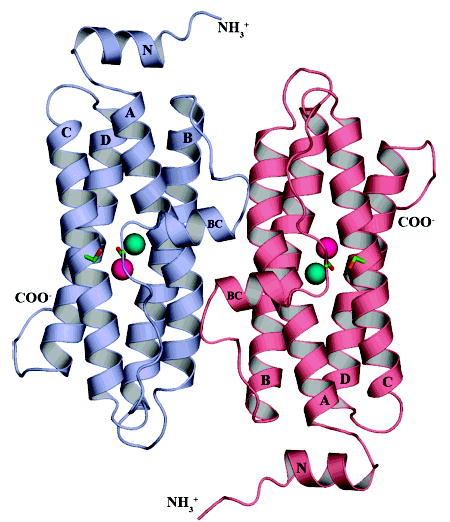
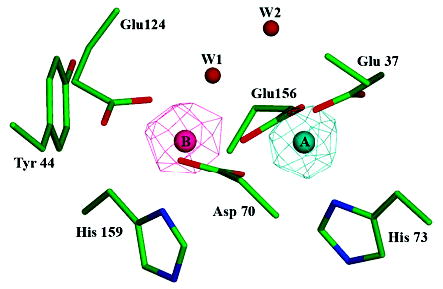
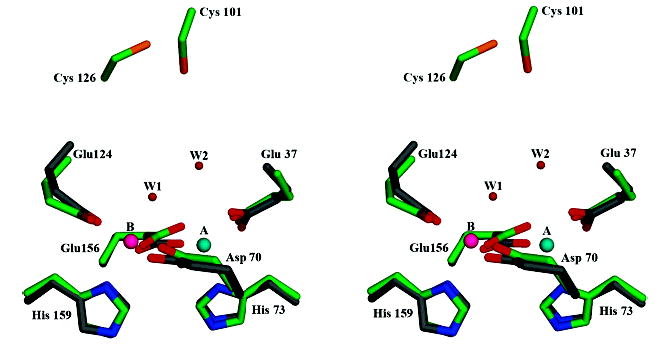
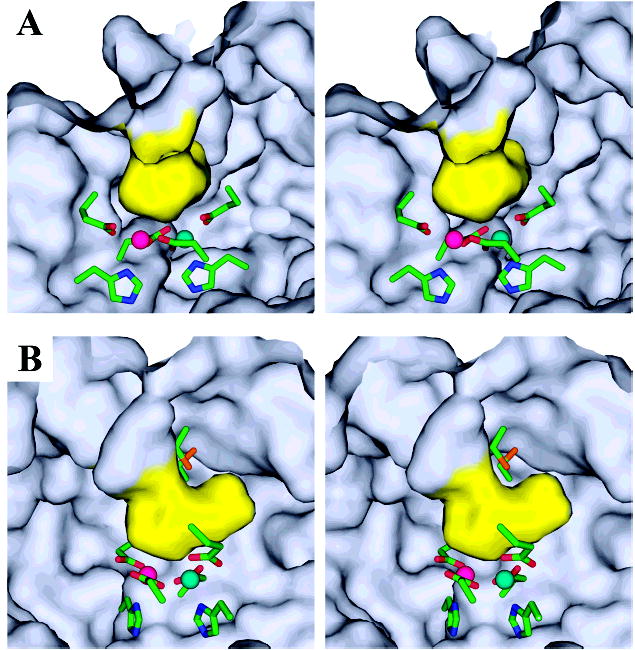
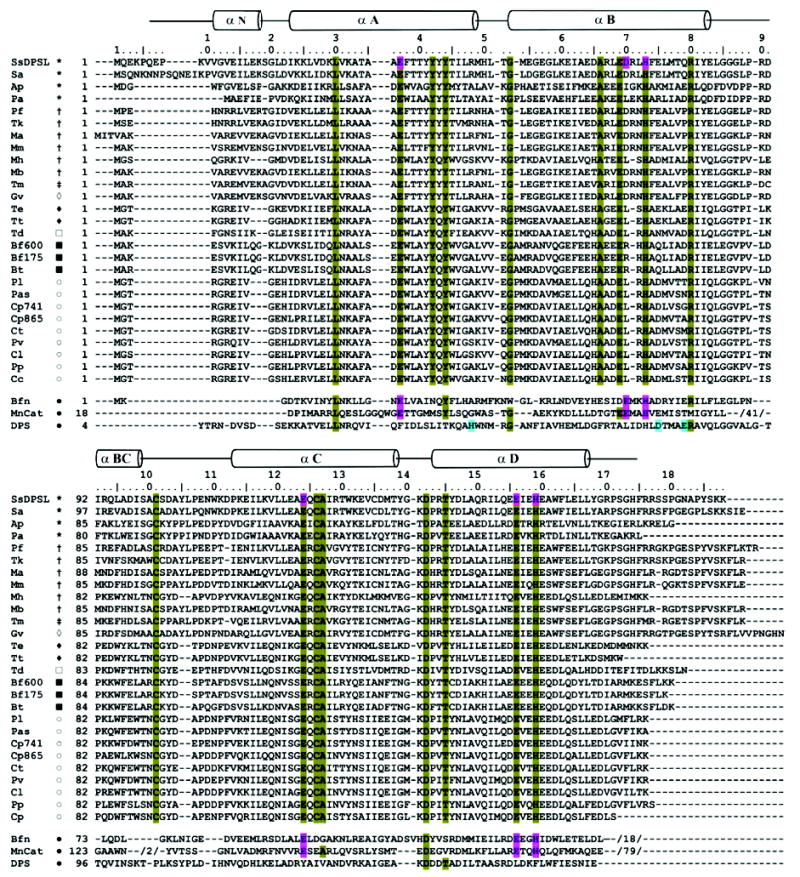
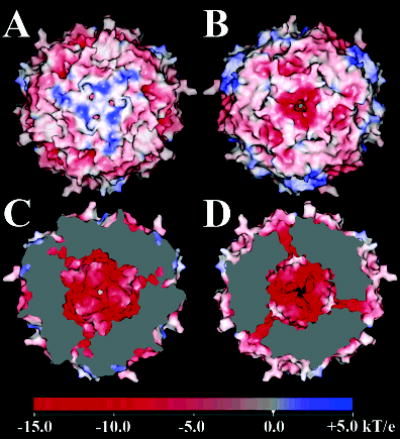
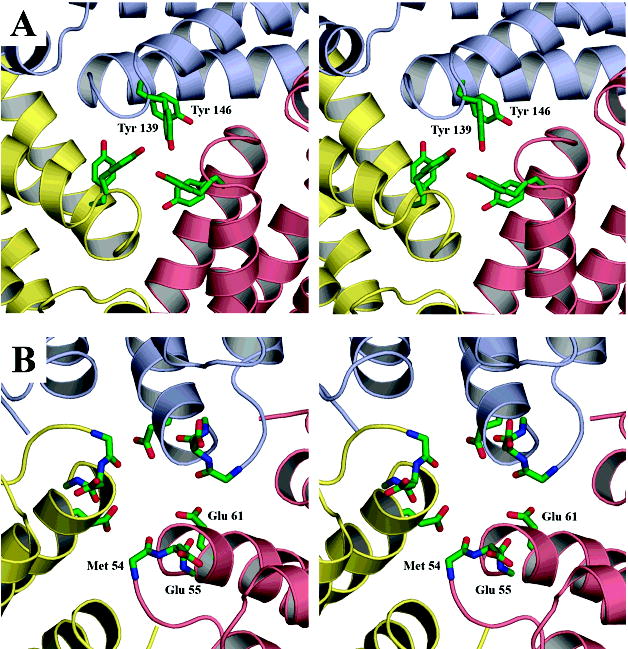
The superfamily of ferritin-like proteins has recently expanded to include a phylogenetically distinct class of proteins termed DPS-like (DPSL) proteins. Despite their distinct genetic signatures, members of this subclass share considerable similarity to previously recognized DPS proteins. Like DPS, these proteins are expressed in response to oxidative stress, form dodecameric cage-like particles, preferentially utilize H(2)O(2) in the controlled oxidation of Fe(2+), and possess a short N-terminal extension implicated in stabilizing cellular DNA. Given these extensive similarities, the functional properties responsible for the preservation of the DPSL signature in the genomes of diverse prokaryotes have been unclear. Here, we describe the crystal structure of a DPSL protein from the thermoacidophilic archaeon Sulfolobus solfataricus. Although the overall fold of the polypeptide chain and the oligomeric state of this protein are indistinguishable from those of authentic DPS proteins, several important differences are observed. First, rather than a ferroxidase site at the subunit interface, as is observed in all other DPS proteins, the ferroxidase site in SsDPSL is buried within the four-helix bundle, similar to bacterioferritin. Second, the structure reveals a channel leading from the exterior surface of SsDPSL to the bacterioferritin-like dimetal binding site, possibly allowing divalent cations and/or H(2)O(2) to access the active site. Third, a pair of cysteine residues unique to DPSL proteins is found adjacent to the dimetal binding site juxtaposed between the exterior surface of the protein and the active site channel. The cysteine residues in this thioferritin motif may play a redox active role, possibly serving to recycle iron at the ferroxidase center.
Figures
References
-
- Andrews SC, Robinson AK, Rodriguez-Quinones F. Bacterial iron homeostasis. FEMS Microbiol Rev. 2003;27:215–237. - PubMed
-
- Baaghil S, Lewin A, Moore GR, Le Brun NE. Core formation in Escherichia coli bacterioferritin requires a functional ferroxidase center. Biochemistry. 2003;42:14047–14056. - PubMed
-
- Zhao GH, Ceci P, Ilari A, Giangiacomo L, Laue TM, Chiancone E, Chasteen ND. Iron and hydrogen peroxide detoxification properties of DNA-binding protein from starved cells: A ferritin-like DNA-binding protein of Escherichia coli. J Biol Chem. 2002;277:27689–27696. - PubMed
-
- Ilari A, Latella MC, Ceci P, Ribacchi F, Su M, Giangiacomo L, Stefanini S, Chasteen ND, Chiancone E. The unusual intersubunit ferroxidase center of Listeria innocua Dps is required for hydrogen peroxide detoxification but not for iron uptake. A study with site-specific mutants. Biochemistry. 2005;44:5579–5587. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
