

NO-independent stimulators and activators of soluble guanylate cyclase: discovery and therapeutic potential
- PMID: 16955067
- PMCID: PMC2225477
- DOI: 10.1038/nrd2038
NO-independent stimulators and activators of soluble guanylate cyclase: discovery and therapeutic potential
Abstract
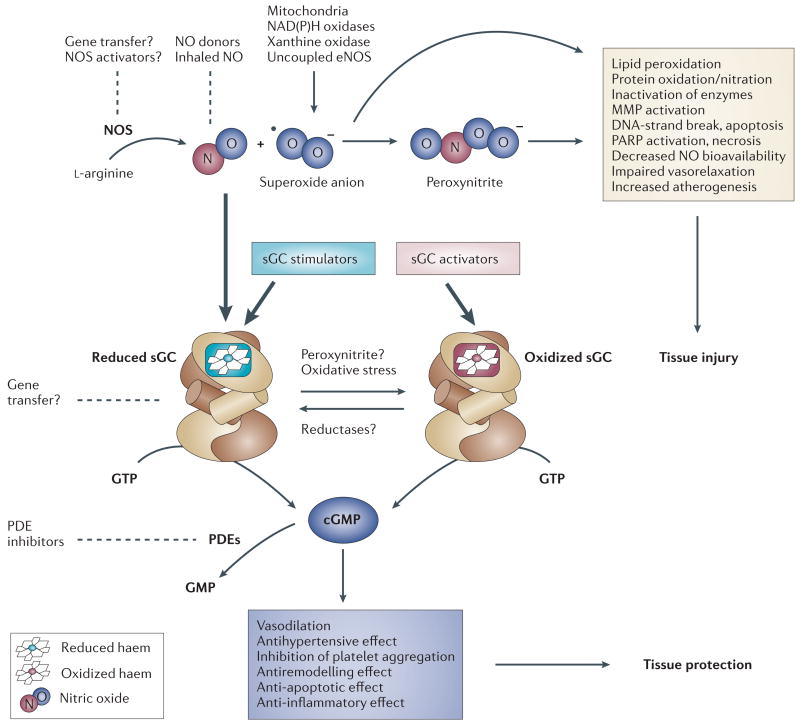
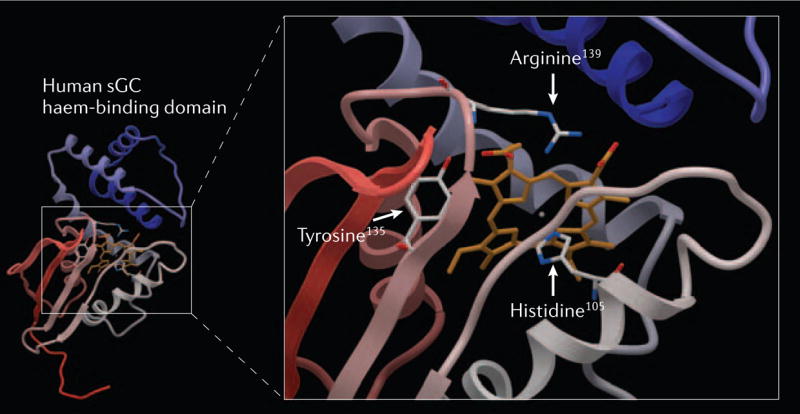
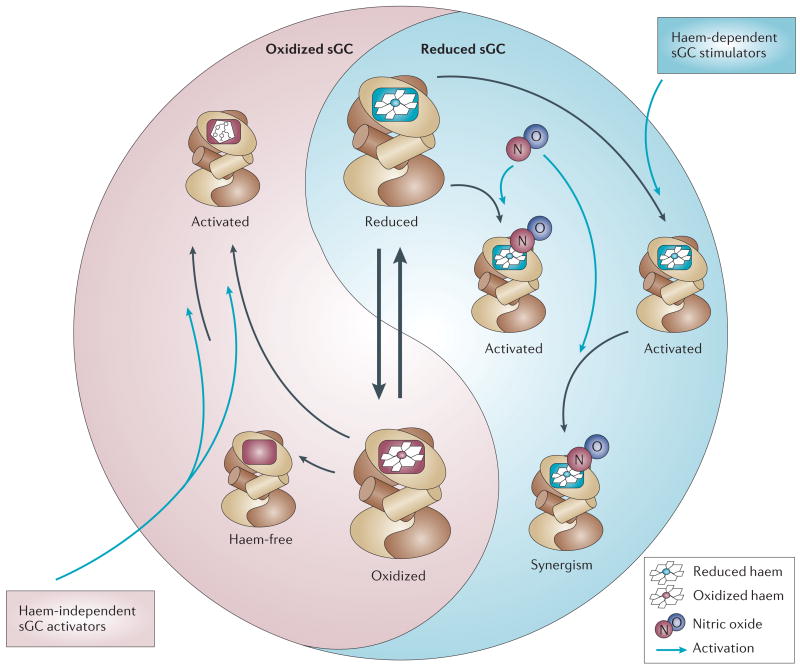
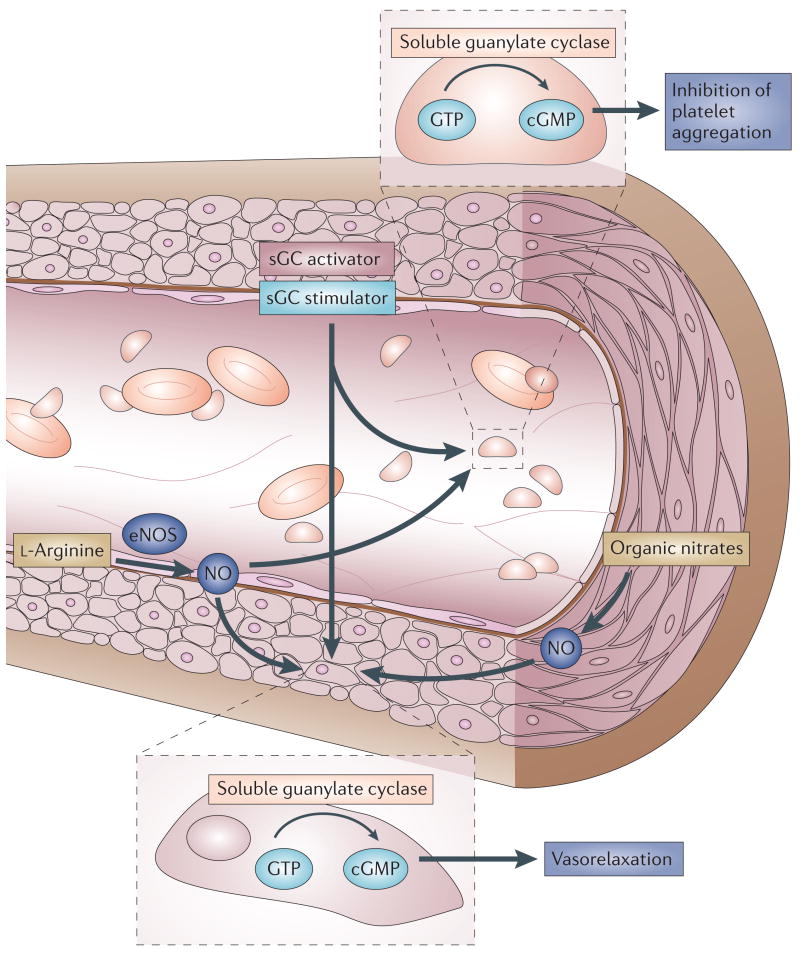
Soluble guanylate cyclase (sGC) is a key signal-transduction enzyme activated by nitric oxide (NO). Impaired bioavailability and/or responsiveness to endogenous NO has been implicated in the pathogenesis of cardiovascular and other diseases. Current therapies that involve the use of organic nitrates and other NO donors have limitations, including non-specific interactions of NO with various biomolecules, lack of response and the development of tolerance following prolonged administration. Compounds that activate sGC in an NO-independent manner might therefore provide considerable therapeutic advantages. Here we review the discovery, biochemistry, pharmacology and clinical potential of haem-dependent sGC stimulators (including YC-1, BAY 41-2272, BAY 41-8543, CFM-1571 and A-350619) and haem-independent sGC activators (including BAY 58-2667 and HMR-1766).
Figures
Similar articles
-
NO-independent, haem-dependent soluble guanylate cyclase stimulators.Handb Exp Pharmacol. 2009;(191):277-308. doi: 10.1007/978-3-540-68964-5_13. Handb Exp Pharmacol. 2009. PMID: 19089334 Review.
-
Exploring the potential of NO-independent stimulators and activators of soluble guanylate cyclase for the medical treatment of erectile dysfunction.Curr Pharm Des. 2010 May;16(14):1619-33. doi: 10.2174/138161210791164162. Curr Pharm Des. 2010. PMID: 20201788 Review.
-
Distinct molecular requirements for activation or stabilization of soluble guanylyl cyclase upon haem oxidation-induced degradation.Br J Pharmacol. 2009 Jul;157(5):781-95. doi: 10.1111/j.1476-5381.2009.00263.x. Epub 2009 May 18. Br J Pharmacol. 2009. PMID: 19466990 Free PMC article.
-
Nitric oxide-independent stimulation of soluble guanylate cyclase with BAY 41-2272 in cardiovascular disease.Cardiovasc Drug Rev. 2007 Spring;25(1):30-45. doi: 10.1111/j.1527-3466.2007.00003.x. Cardiovasc Drug Rev. 2007. PMID: 17445086 Review.
-
Molecular mechanisms underlying rat mesenteric artery vasorelaxation induced by the nitric oxide-independent soluble guanylyl cyclase stimulators BAY 41-2272 [5-cyclopropyl-2-[1-(2-fluorobenzyl)-1H-pyrazolo[3,4-b]pyridin-3-yl]pyrimidin-4-ylamine] and YC-1 [3-(5'-hydroxymethyl-2'-furyl)-1-benzyl Indazole].J Pharmacol Exp Ther. 2006 Apr;317(1):258-66. doi: 10.1124/jpet.105.095752. Epub 2005 Dec 13. J Pharmacol Exp Ther. 2006. PMID: 16352702
Cited by
-
Emerging Pharmacologic Therapies for Heart Failure With Reduced Ejection Fraction.CJC Open. 2021 Feb 1;3(5):646-657. doi: 10.1016/j.cjco.2021.01.006. eCollection 2021 May. CJC Open. 2021. PMID: 34027369 Free PMC article. Review.
-
Novel vasodilators in heart failure.Curr Heart Fail Rep. 2013 Mar;10(1):1-11. doi: 10.1007/s11897-012-0126-4. Curr Heart Fail Rep. 2013. PMID: 23299783 Review.
-
Cyclic guanosine monophosphate signaling and phosphodiesterase-5 inhibitors in cardioprotection.J Am Coll Cardiol. 2012 May 29;59(22):1921-7. doi: 10.1016/j.jacc.2011.09.086. J Am Coll Cardiol. 2012. PMID: 22624832 Free PMC article. Review.
-
Vericiguat in Heart Failure: Characteristics, Scientific Evidence and Potential Clinical Applications.Biomedicines. 2022 Oct 3;10(10):2471. doi: 10.3390/biomedicines10102471. Biomedicines. 2022. PMID: 36289733 Free PMC article. Review.
-
Effects of age and sex on the pharmacokinetics of the soluble guanylate cyclase stimulator riociguat (BAY 63-2521).Pulm Circ. 2016 Mar;6(Suppl 1):S58-65. doi: 10.1086/685019. Pulm Circ. 2016. PMID: 27162629 Free PMC article.
References
-
- Feelisch M. The use of nitric oxide donors in pharmacological studies. Naunyn Schmiedebergs Arch Pharmacol. 1998;358:113–122. - PubMed
-
- Munzel T, Daiber A, Mulsch A. Explaining the phenomenon of nitrate tolerance. Circ Res. 2005;97:618–628. - PubMed
-
- Dikalov S, Fink B, Skatchkov M, Stalleicken D, Bassenge E. Formation of reactive oxygen species by pentaerithrityltetranitrate and glyceryl trinitrate in vitro and development of nitrate tolerance. J Pharmacol Exp Ther. 1998;286:938–944. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
