

Multiple contributions of peroxisomal metabolic function to fungal pathogenicity in Colletotrichum lagenarium
- PMID: 16957261
- PMCID: PMC1563638
- DOI: 10.1128/AEM.00988-06
Multiple contributions of peroxisomal metabolic function to fungal pathogenicity in Colletotrichum lagenarium
Abstract
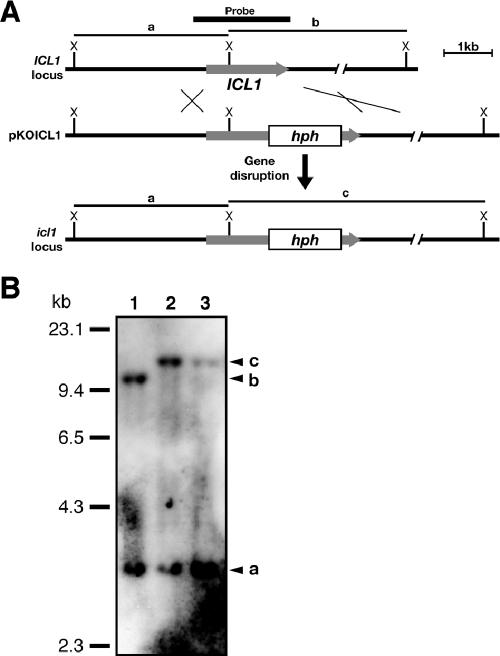
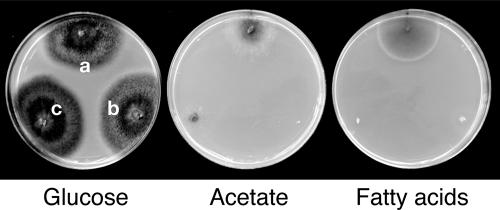
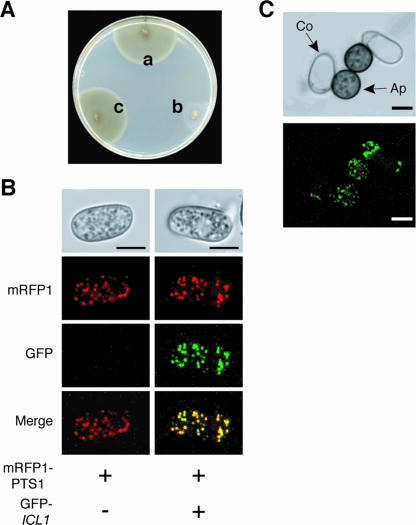
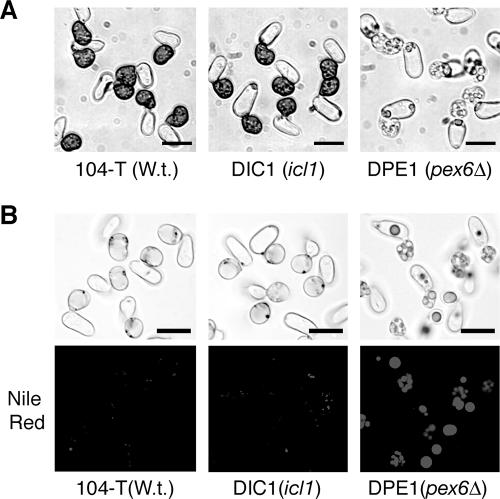
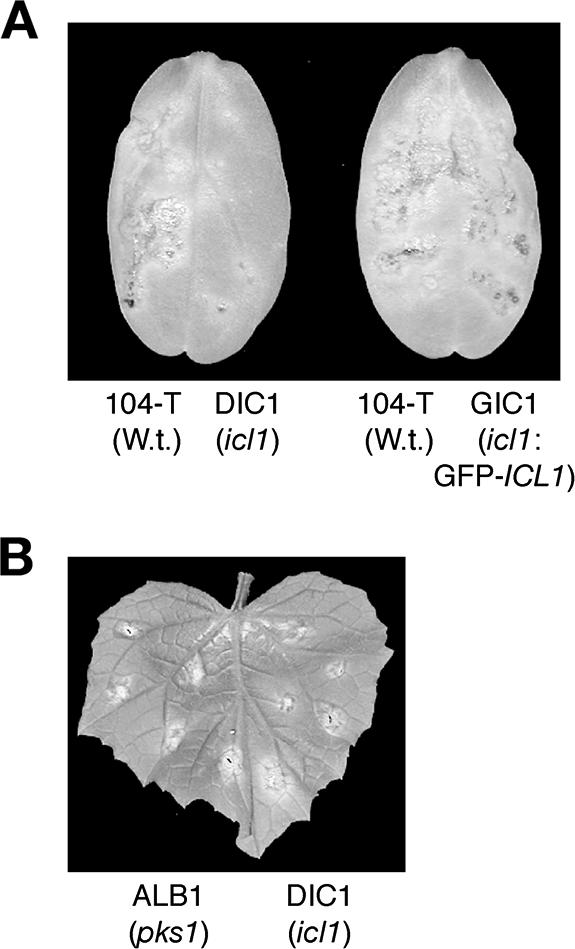
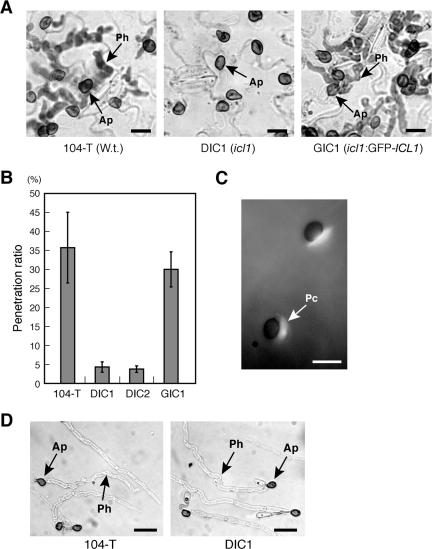
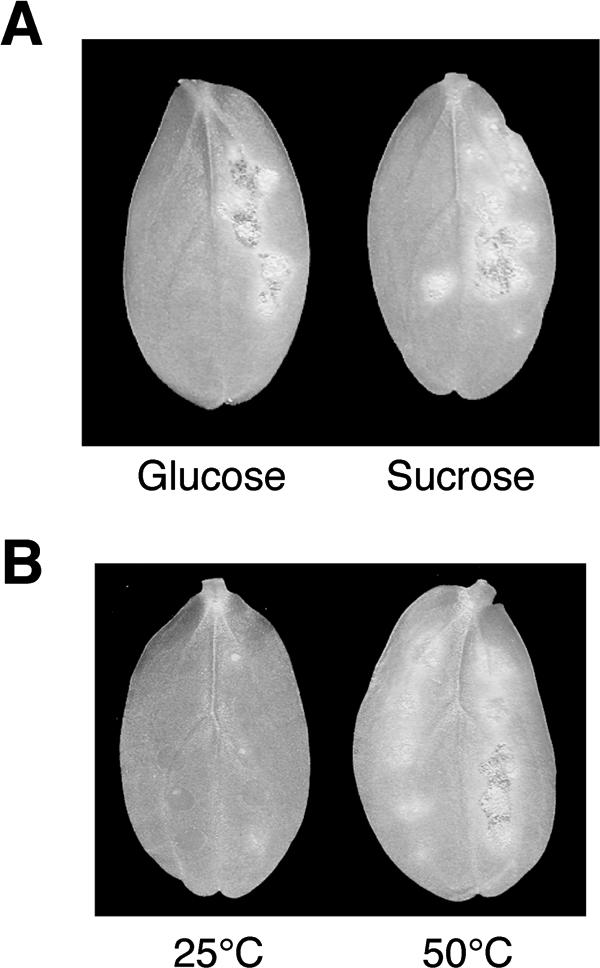
In Colletotrichum lagenarium, which is the causal agent of cucumber anthracnose, PEX6 is required for peroxisome biogenesis and appressorium-mediated infection. To verify the roles of peroxisome-associated metabolism in fungal pathogenicity, we isolated and functionally characterized ICL1 of C. lagenarium, which encodes isocitrate lyase involved in the glyoxylate cycle in peroxisomes. The icl1 mutants failed to utilize fatty acids and acetate for growth. Although Icl1 has no typical peroxisomal targeting signals, expression analysis of the GFP-Icl1 fusion protein indicated that Icl1 localizes in peroxisomes. These results indicate that the glyoxylate cycle that occurs inside the peroxisome is required for fatty acid and acetate metabolism for growth. Importantly, in contrast with the pex6 mutants that form nonmelanized appressoria, the icl1 mutants formed appressoria that were highly pigmented with melanin, suggesting that the glyoxylate cycle is not essential for melanin biosynthesis in appressoria. However, the icl1 mutants exhibited a severe reduction in virulence. Appressoria of the icl1 mutants failed to develop penetration hyphae in the host plant, suggesting that ICL1 is involved in host invasion. The addition of glucose partially restored virulence of the icl1 mutant. Heat shock treatment of the host plant also enabled the icl1 mutants to develop lesions, implying that the infection defect of the icl1 mutant is associated with plant defense. Together with the requirement of PEX6 for appressorial melanization, our findings suggest that peroxisomal metabolic pathways play functional roles in appressorial melanization and subsequent host invasion steps, and the latter step requires the glyoxylate cycle.
Figures

References
-
- Adam, L., and S. C. Somerville. 1996. Genetic characterization of five powdery mildew disease resistance loci in Arabidopsis thaliana. Plant J. 9:341-356. - PubMed
-
- Agrios, G. N. 2004. Plant pathology, 5th ed., p. 487-498. Academic Press, San Diego, Calif.
-
- Emmett, R. W., and D. G. Parbery. 1975. Appressoria. Annu. Rev. Phytopathol. 13:147-167.
-
- Fujii, I., Y. Mori, A. Watanabe, Y. Kubo, G. Tsuji, and Y. Ebizuka. 2000. Enzymatic synthesis of 1,3,6,8-tetrahydroxynaphthalene solely from malonyl coenzyme A by a fungal iterative type I polyketide synthase PKS1. Biochemistry 39:8853-8858. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
