

Distinct roles of doublecortin modulating the microtubule cytoskeleton
- PMID: 16957770
- PMCID: PMC1590004
- DOI: 10.1038/sj.emboj.7601335
Distinct roles of doublecortin modulating the microtubule cytoskeleton
Abstract
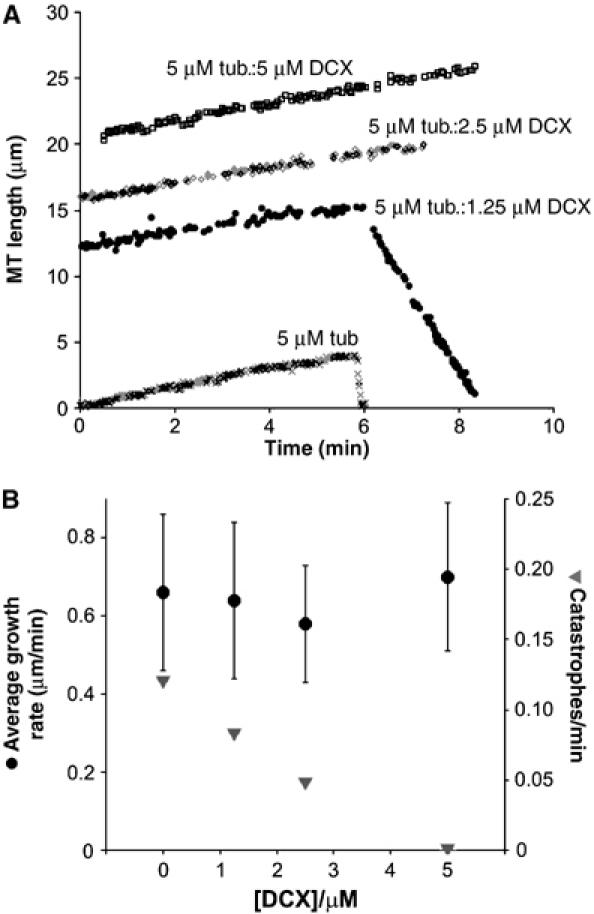
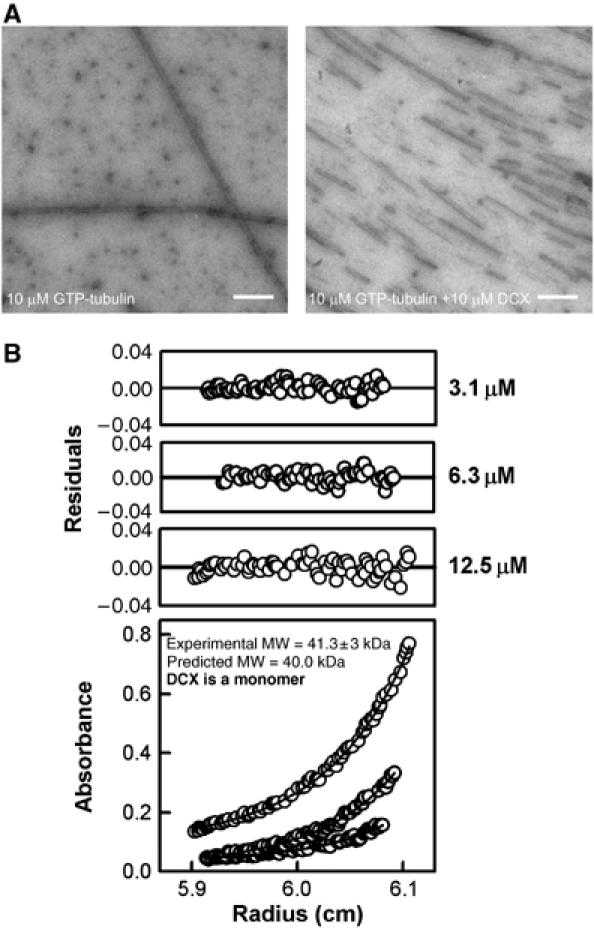
Doublecortin is a neuronal microtubule-stabilising protein, mutations of which cause mental retardation and epilepsy in humans. How doublecortin influences microtubule dynamics, and thereby brain development, is unclear. We show here by video microscopy that purified doublecortin has no effect on the growth rate of microtubules. However, it is a potent anti-catastrophe factor that stabilises microtubules by linking adjacent protofilaments and counteracting their outward bending in depolymerising microtubules. We show that doublecortin-stabilised microtubules are substrates for kinesin translocase motors and for depolymerase kinesins. In addition, doublecortin does not itself oligomerise and does not bind to tubulin heterodimers but does nucleate microtubules. In cells, doublecortin is enriched at the distal ends of neuronal processes and our data raise the possibility that the function of doublecortin in neurons is to drive assembly and stabilisation of non-centrosomal microtubules in these doublecortin-enriched distal zones. These distinct properties combine to give doublecortin a unique function in microtubule regulation, a role that cannot be compensated for by other microtubule-stabilising proteins and nucleating factors.
Figures




References
-
- Ackmann M, Wiech H, Mandelkow E (2000) Nonsaturable binding indicates clustering of tau on the microtubule surface in a paired helical filament-like conformation. J Biol Chem 275: 30335–30343 - PubMed
-
- Aldaz H, Rice LM, Stearns T, Agard DA (2005) Insights into microtubule nucleation from the crystal structure of human γ-tubulin. Nature 435: 523–527 - PubMed
-
- Andersen SSL (2005) The search and prime hypothesis for growth cone turning. BioEssays 27: 86–90 - PubMed
-
- Arnal I, Heichette C, Diamantopoulos GS, Chretien D (2004) CLIP-170/tubulin-curved oligomers coassemble at microtubule ends and promote rescues. Curr Biol 14: 2086–2095 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
