

Silencing by plant Polycomb-group genes requires dispersed trimethylation of histone H3 at lysine 27
- PMID: 16957776
- PMCID: PMC1590001
- DOI: 10.1038/sj.emboj.7601311
Silencing by plant Polycomb-group genes requires dispersed trimethylation of histone H3 at lysine 27
Abstract
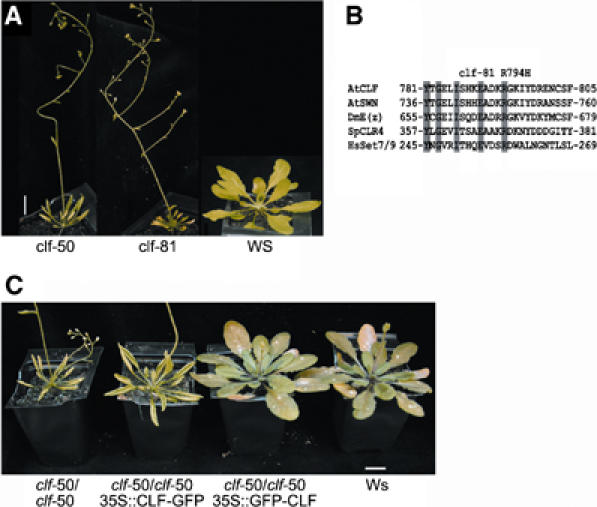
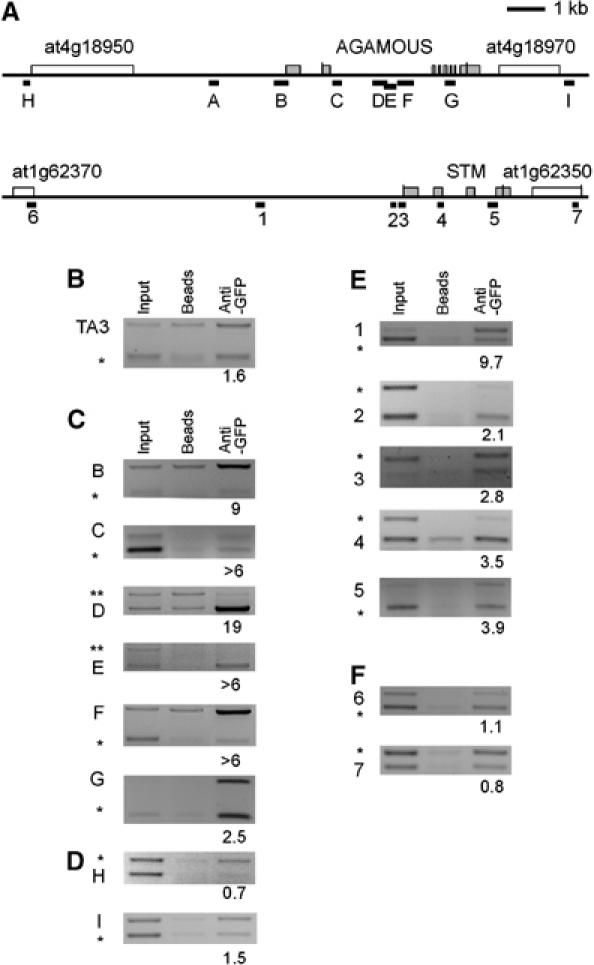
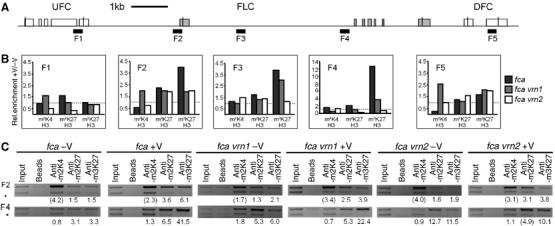
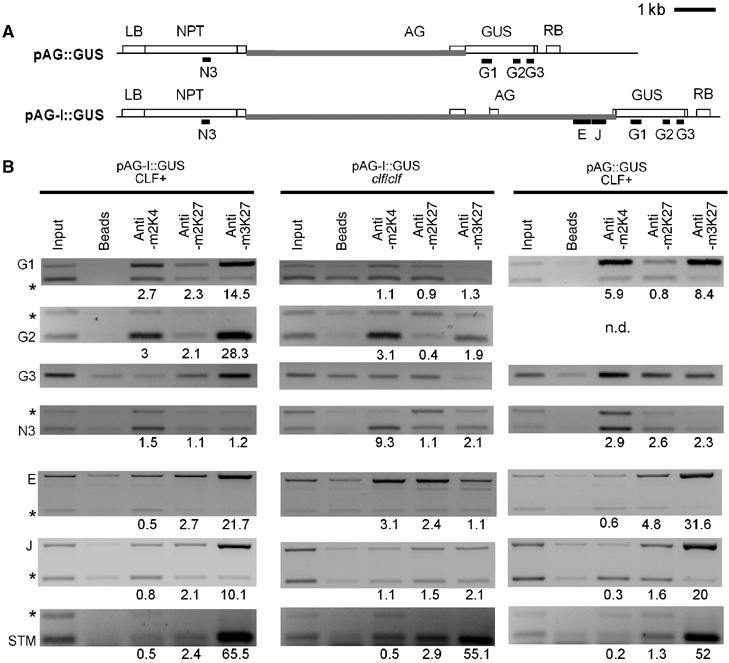
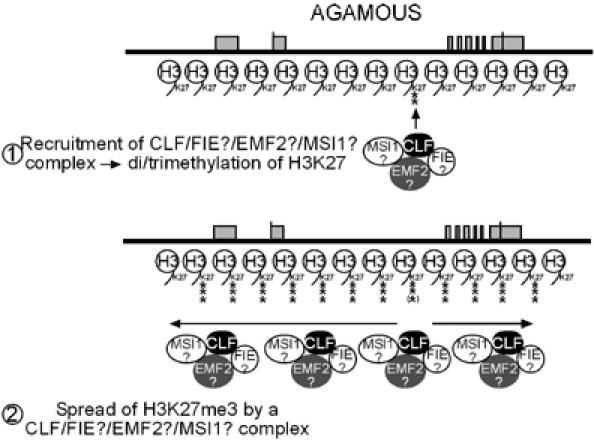
The plant Polycomb-group (Pc-G) protein CURLY LEAF (CLF) is required to repress targets such as AGAMOUS (AG) and SHOOTMERISTEMLESS (STM). Using chromatin immunoprecipitation, we identify AG and STM as direct targets for CLF and show that they carry a characteristic epigenetic signature of dispersed histone H3 lysine 27 trimethylation (H3K27me3) and localised H3K27me2 methylation. H3K27 methylation is present throughout leaf development and consistent with this, CLF is required persistently to silence AG. However, CLF is not itself an epigenetic mark as it is lost during mitosis. We suggest a model in which Pc-G proteins are recruited to localised regions of targets and then mediate dispersed H3K27me3. Analysis of transgenes carrying AG regulatory sequences confirms that H3K27me3 can spread to novel sequences in a CLF-dependent manner and further shows that H3K27me3 methylation is not sufficient for silencing of targets. We suggest that the spread of H3K27me3 contributes to the mitotic heritability of Pc-G silencing, and that the loss of silencing caused by transposon insertions at plant Pc-G targets reflects impaired spreading.
Figures


References
-
- Annunziato AT (2005) Split decision: what happens to nucleosomes during DNA replication? J Biol Chem 280: 12065–12068 - PubMed
-
- Bannister AJ, Schneider R, Kouzarides T (2002) Histone methylation: dynamic or static? Cell 109: 801–806 - PubMed
-
- Bastow R, Mylne JS, Lister C, Lippman Z, Martienssen RA, Dean C (2004) Vernalization requires epigenetic silencing of FLC by histone methylation. Nature 427: 164–167 - PubMed
-
- Baumbusch LO, Thorstensen T, Krauss V, Fischer A, Naumann K, Assalkhou R, Schulz I, Reuter G, Aalen RB (2001) The Arabidopsis thaliana genome contains at least 29 active genes encoding SET domain proteins that can be assigned to four evolutionarily conserved classes. Nucleic Acids Res 29: 4319–4333 - PMC - PubMed
-
- Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, Fry B, Meissner A, Wernig M, Plath K, Jaenisch R, Wagschal A, Feil R, Schreiber SL, Lander ES (2006) A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell 125: 315–326 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
