

Forespore engulfment mediated by a ratchet-like mechanism
- PMID: 16959571
- PMCID: PMC3266857
- DOI: 10.1016/j.cell.2006.06.053
Forespore engulfment mediated by a ratchet-like mechanism
Abstract
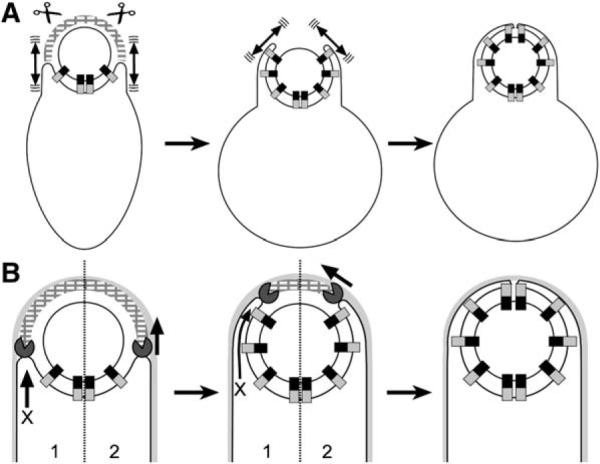
A key step in bacterial endospore formation is engulfment, during which one bacterial cell engulfs another in a phagocytosis-like process that normally requires SpoIID, SpoIIM, and SpoIIP (DMP). We here describe a second mechanism involving the zipper-like interaction between the forespore protein SpoIIQ and its mother cell ligand SpoIIIAH, which are essential for engulfment when DMP activity is reduced or SpoIIB is absent. They are also required for the rapid engulfment observed during the enzymatic removal of peptidoglycan, a process that does not require DMP. These results suggest the existence of two separate engulfment machineries that compensate for one another in intact cells, thereby rendering engulfment robust. Photobleaching analysis demonstrates that SpoIIQ assembles a stationary structure, suggesting that SpoIIQ and SpoIIIAH function as a ratchet that renders forward membrane movement irreversible. We suggest that ratchet-mediated engulfment minimizes the utilization of chemical energy during this dramatic cellular reorganization, which occurs during starvation.
Figures





References
-
- Antal T, Krapivsky PL. “Burnt-bridge” mechanism of molecular motor motion. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 2005 in press. Published online October 6, 2005. 10.1103/PhysRevE.72.046104. - PubMed
-
- Bar-Nahum G, Epshtein V, Ruckenstein AE, Rafikov R, Mustaev A, Nudler E. A ratchet mechanism of transcription elongation and its control. Cell. 2005;120:183–193. - PubMed
-
- Cutting S, Roels S, Losick R. Sporulation operon spoIVF and the characterization of mutations that uncouple mother-cell from forespore gene expression in Bacillus subtilis. J. Mol. Biol. 1991;221:1237–1256. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
