

Distinct contributions of vaccine-induced immunoglobulin G1 (IgG1) and IgG2a antibodies to protective immunity against influenza
- PMID: 16960108
- PMCID: PMC1563571
- DOI: 10.1128/CVI.00156-06
Distinct contributions of vaccine-induced immunoglobulin G1 (IgG1) and IgG2a antibodies to protective immunity against influenza
Abstract
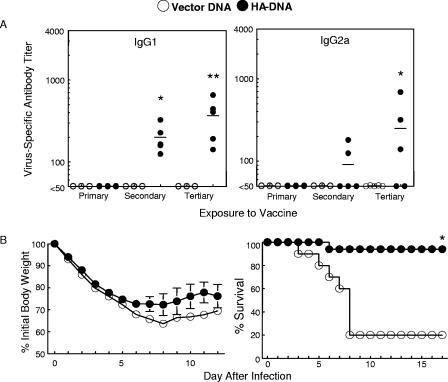
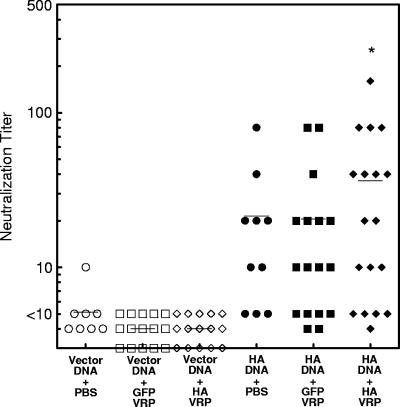
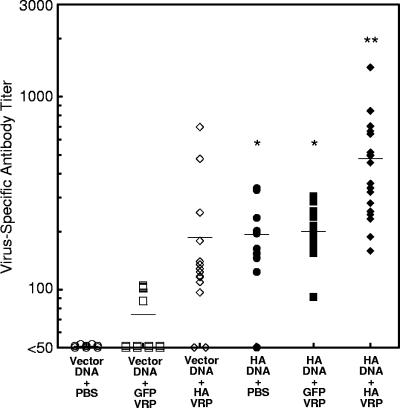
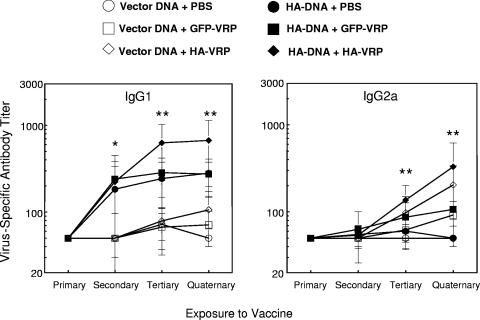
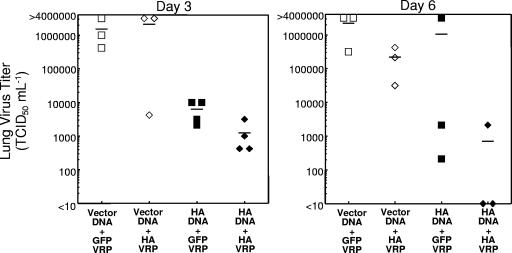
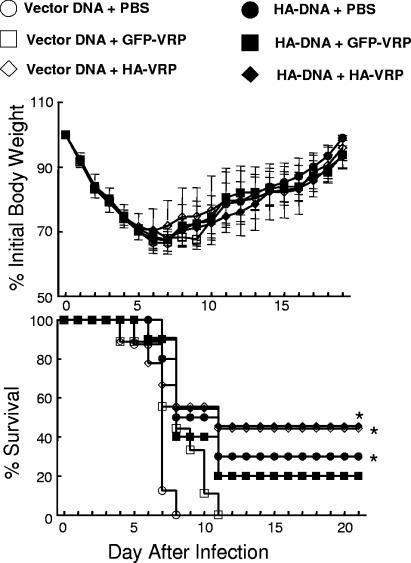
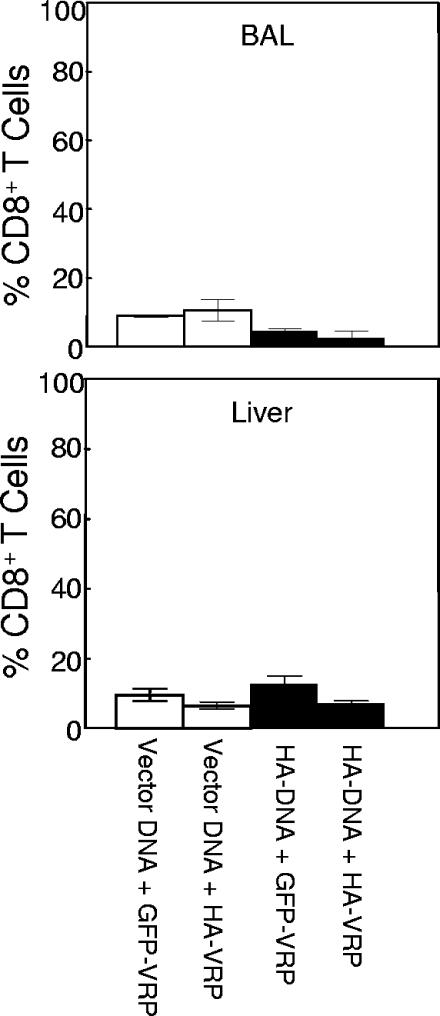
Vaccination represents the most effective form of protection against influenza infection. While neutralizing antibodies are typically measured as a correlate of vaccine-induced protective immunity against influenza, nonneutralizing antibodies may contribute to protection or amelioration of disease. The goal of this study was to dissect the individual contributions of the immunoglobulin G1 (IgG1) and IgG2a antibody isotypes to vaccine-induced immunity against influenza virus. To accomplish this, we utilized an influenza vaccine regimen that selectively enhanced IgG1 or IgG2a antibodies by using either DNA or viral replicon particle (VRP) vectors expressing influenza virus hemagglutinin (HA) (HA-DNA or HA-VRP, respectively). After HA-DNA vaccination, neutralizing antibodies were detected by both in vitro (microneutralization) and in vivo (lung viral titer) methods and were associated with increased IgG1 expression by enzyme-linked immunosorbent assay (ELISA). Vaccination with HA-VRP did not strongly stimulate either neutralizing or IgG1 antibodies but did induce IgG2a antibodies. Expression of IgG2a antibodies in this context correlated with clearance of virus and increased protection against lethal influenza challenge. Increased induction of both antibody isotypes as measured by ELISA was a better correlate for vaccine efficacy than neutralization alone. This study details separate but important roles for both IgG1 and IgG2a expression in vaccination against influenza and argues for the development of vaccine regimens that stimulate and measure expression of both antibody isotypes.
Figures
References
-
- Arulanandam, B. P., M. O'Toole, and D. W. Metzger. 1999. Intranasal interleukin-12 is a powerful adjuvant for protective mucosal immunity. J. Infect. Dis. 180:940-949. - PubMed
-
- Barackman, J. D., G. Ott, and D. T. O'Hagan. 1999. Intranasal immunization of mice with influenza vaccine in combination with the adjuvant LT-R72 induces potent mucosal and serum immunity which is stronger than that with traditional intramuscular immunization. Infect. Immun. 67:4276-4279. - PMC - PubMed
-
- Beigel, J. H., J. Farrar, A. M. Han, F. G. Hayden, R. Hyer, M. D. de Jong, S. Lochindarat, T. K. Nguyen, T. H. Nguyen, T. H. Tran, A. Nicoll, S. Touch, K. Y. Yuen, et al. 2005. Avian influenza A (H5N1) infection in humans. N. Engl. J. Med. 353:1374-1385. - PubMed
-
- Belshe, R. B., W. C. Gruber, P. M. Mendelman, H. B. Mehta, K. Mahmood, K. Reisinger, J. Treanor, K. Zangwill, F. G. Hayden, D. I. Bernstein, K. Kotloff, J. King, P. A. Piedra, S. L. Block, L. Yan, and M. Wolff. 2000. Correlates of immune protection induced by live, attenuated, cold-adapted, trivalent, intranasal influenza virus vaccine. J. Infect. Dis. 181:1133-1137. - PubMed
-
- Benne, C. A., M. Harmsen, W. van der Graaff, A. F. Verheul, H. Snippe, and C. A. Kraaijeveld. 1997. Influenza virus neutralizing antibodies and IgG isotype profiles after immunization of mice with influenza A subunit vaccine using various adjuvants. Vaccine 15:1039-1044. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
