

Dynamic and static calcium gradients inside large snail (Helix aspersa) neurones detected with calcium-sensitive microelectrodes
- PMID: 16962659
- PMCID: PMC1885971
- DOI: 10.1016/j.ceca.2006.07.010
Dynamic and static calcium gradients inside large snail (Helix aspersa) neurones detected with calcium-sensitive microelectrodes
Abstract
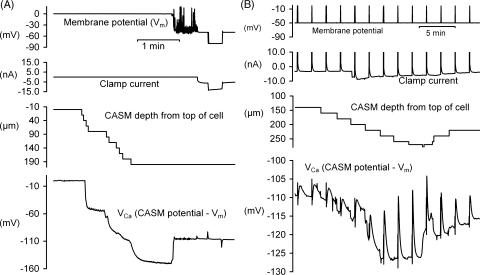
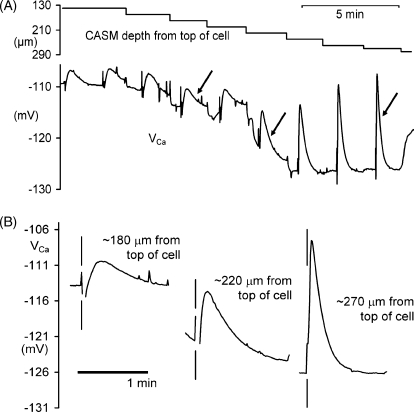
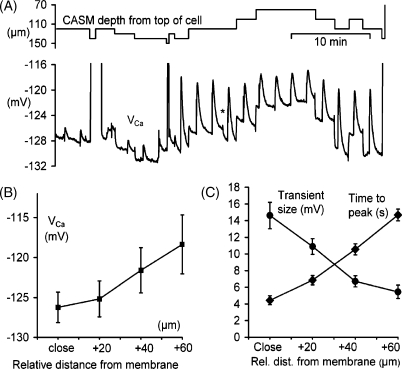
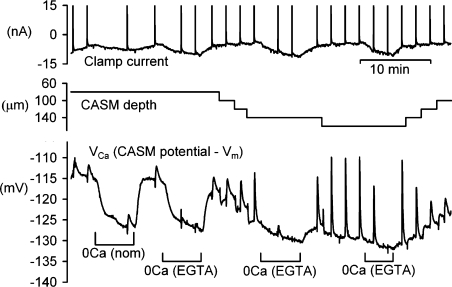
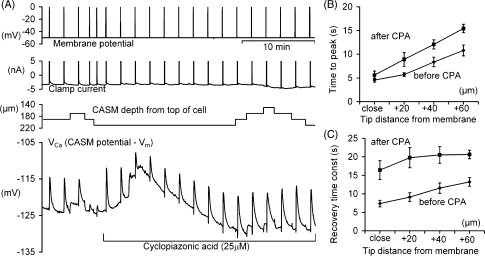
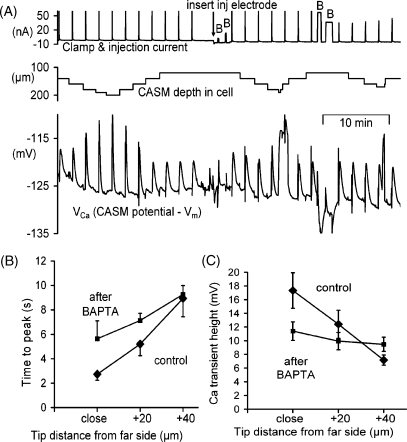
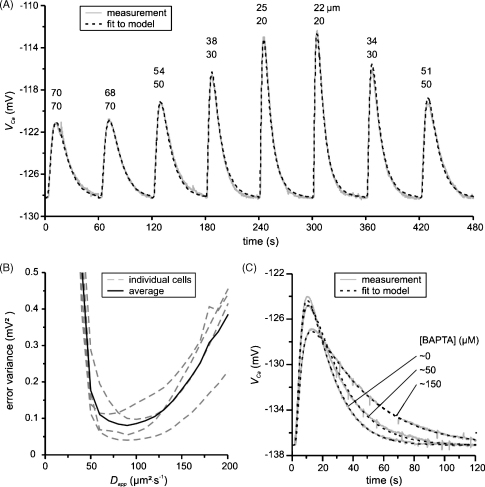
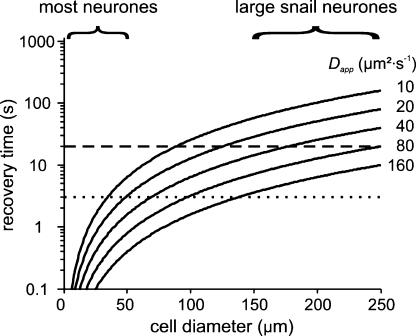
We have used quartz Ca2+-sensitive microelectrodes (CASMs) in large voltage-clamped snail neurones to investigate the inward spread of Ca2+ after a brief depolarisation. Both steady state and [Ca2+]i transients changed with depth of penetration. When the CASM tip was within 20 microm of the far side of the cell the [Ca2+]i transient time to peak was 4.4+/-0.5s, rising to 14.7+/-0.7s at a distance of 80 microm. We estimate that the Ca2+ transients travelled centripetally at an average speed of 6 microm2 s(-1) and decreased in size by half over a distance of about 45 microm. Cyclopiazonic acid had little effect on the size and time to peak of Ca2+ transients but slowed their recovery significantly. This suggests that the endoplasmic reticulum curtails rather than reinforces the transients. Injecting the calcium buffer BAPTA made the Ca2+ transients more uniform in size and increased their times to peak and rates of recovery near the membrane. We have developed a computational model for the transients, which includes diffusion, uptake and Ca2+ extrusion. Good fits were obtained with a rather large apparent diffusion coefficient of about 90+/-20 microm2 s(-1). This may assist fast recovery by extrusion.
Figures
Similar articles
-
Relationship between intracellular calcium and its muffling measured by calcium iontophoresis in snail neurones.J Physiol. 1996 Mar 15;491 ( Pt 3)(Pt 3):621-33. doi: 10.1113/jphysiol.1996.sp021244. J Physiol. 1996. PMID: 8815198 Free PMC article.
-
Effects of injecting calcium-buffer solution on [Ca2+]i in voltage-clamped snail neurons.Biophys J. 1996 May;70(5):2120-30. doi: 10.1016/S0006-3495(96)79778-3. Biophys J. 1996. PMID: 9172736 Free PMC article.
-
Calcium clamp in isolated neurones of the snail Helix pomatia.J Physiol. 1993 Mar;462:47-58. doi: 10.1113/jphysiol.1993.sp019542. J Physiol. 1993. PMID: 8392572 Free PMC article.
-
Ability of the Ca2+-selective microelectrodes to measure fast and local Ca2+ transients in nerve cells.Can J Physiol Pharmacol. 1987 May;65(5):904-14. doi: 10.1139/y87-146. Can J Physiol Pharmacol. 1987. PMID: 2441832 Review.
-
The role of calcium in cell division.Cell Calcium. 1994 Oct;16(4):322-30. doi: 10.1016/0143-4160(94)90096-5. Cell Calcium. 1994. PMID: 7820852 Review.
Cited by
-
The Ca(2+): H(+) coupling ratio of the plasma membrane calcium ATPase in neurones is little sensitive to changes in external or internal pH.Cell Calcium. 2011 Jun;49(6):357-64. doi: 10.1016/j.ceca.2011.03.004. Epub 2011 Apr 3. Cell Calcium. 2011. PMID: 21466891 Free PMC article.
-
Modeling of the modulation by buffers of Ca2+ release through clusters of IP3 receptors.Biophys J. 2009 Aug 19;97(4):992-1002. doi: 10.1016/j.bpj.2009.05.050. Biophys J. 2009. PMID: 19686646 Free PMC article.
-
The plasma membrane calcium ATPase (PMCA) of neurones is electroneutral and exchanges 2 H+ for each Ca2+ or Ba2+ ion extruded.J Physiol. 2009 Jan 15;587(2):315-27. doi: 10.1113/jphysiol.2008.162453. Epub 2008 Dec 8. J Physiol. 2009. PMID: 19064619 Free PMC article.
References
-
- Berridge M.J. Neuronal calcium signalling. Neuron. 1998;21:13–26. - PubMed
-
- Petersen O.H., Michalak M., Verkhratsky A. Calcium signalling: past, present and future. Cell Calcium. 2005;38:161–169. - PubMed
-
- Fields R.D., Lee P.R., Cohen J.E. Temporal integration of intracellular Ca2+ signaling networks in regulating gene expression by action potentials. Cell Calcium. 2005;37:433–442. - PubMed
-
- Marius P., Guerra M.T., Nathanson M.H., Ehrlich B.E., Leite M.F. Calcium release from ryanodine receptors in the nucleoplasmic reticulum. Cell Calcium. 2006;39:65–73. - PubMed
-
- Grynkiewicz G., Poenie M., Tsien R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985;260:3440–3450. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
