

Direct interaction between caldesmon and cortactin
- PMID: 16962992
- PMCID: PMC1847647
- DOI: 10.1016/j.abb.2006.07.018
Direct interaction between caldesmon and cortactin
Abstract
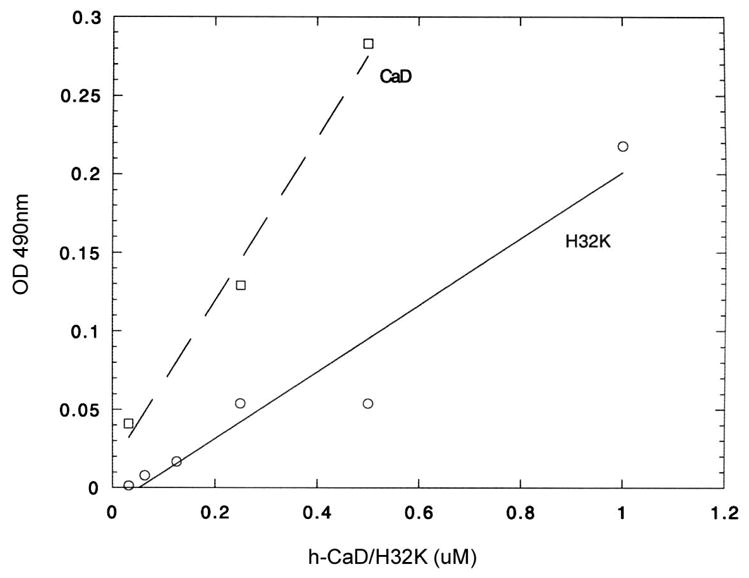
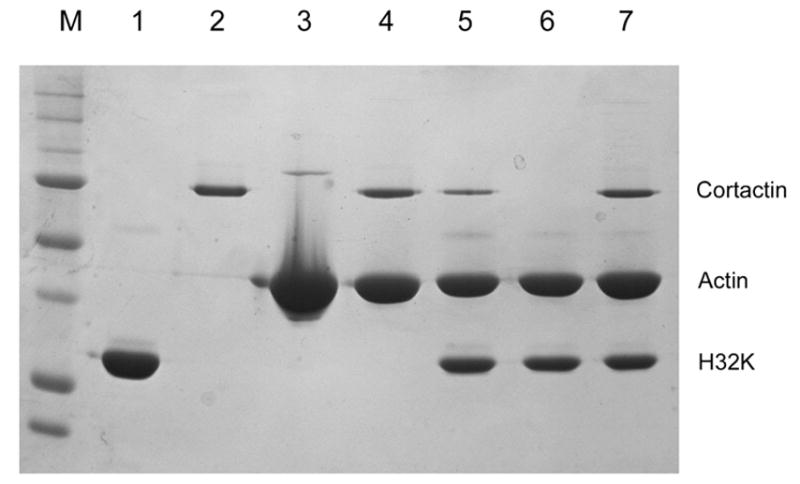
Actin polymerization and depolymerization plays a central role in controlling a wide spectrum of cellular processes. There are many actin-binding proteins in eukaryotic cells. Their roles in the remodeling of the actin architecture and whether they work cooperatively await further study. Caldesmon (CaD) is an actin-binding protein present in nearly all mammalian cells. Cortactin is another actin-binding protein found mainly in the cell cortex. There have been no reports suggesting that CaD and cortactin interact with each other or work as partners. Here, we present evidence that CaD binds cortactin directly by overlay, pull-down assays, ELISA, and by column chromatography. The interaction involves the N-terminal region of cortactin and the C-terminal region of CaD, and appears to be enhanced by divalent metal ions. Cortactin competes with both full-length CaD and its C-terminal fragment for actin binding. Binding of cortactin partially alleviates the inhibitory effect of CaD on the actomyosin ATPase activity. Not only can binding be demonstrated in vitro, the two proteins also co-localize in activated cells at the cortex. Whether such interactions bear any functional significance awaits further investigation.
Figures






Similar articles
-
Caldesmon binding to actin is regulated by calmodulin and phosphorylation via different mechanisms.Biochemistry. 2003 Mar 11;42(9):2513-23. doi: 10.1021/bi0268605. Biochemistry. 2003. PMID: 12614145
-
Structure-activity studies of the regulatory interaction of the 10 kilodalton C-terminal fragment of caldesmon with actin and the effect of mutation of caldesmon residues 691-696.Biochemistry. 1998 Feb 24;37(8):2314-26. doi: 10.1021/bi971727b. Biochemistry. 1998. PMID: 9485378
-
Role of the basic C-terminal half of caldesmon in its regulation of F-actin: comparison between caldesmon and calponin.J Biochem. 2005 Dec;138(6):805-13. doi: 10.1093/jb/mvi181. J Biochem. 2005. PMID: 16428310
-
Phosphorylation of caldesmon during smooth muscle contraction and cell migration or proliferation.J Biomed Sci. 2006 Mar;13(2):159-72. doi: 10.1007/s11373-005-9060-8. J Biomed Sci. 2006. PMID: 16453176 Review.
-
Cortactin branches out: roles in regulating protrusive actin dynamics.Cell Motil Cytoskeleton. 2008 Sep;65(9):687-707. doi: 10.1002/cm.20296. Cell Motil Cytoskeleton. 2008. PMID: 18615630 Free PMC article. Review.
Cited by
-
Comprehensive Proteomic and Metabolomic Signatures of Nontypeable Haemophilus influenzae-Induced Acute Otitis Media Reveal Bacterial Aerobic Respiration in an Immunosuppressed Environment.Mol Cell Proteomics. 2016 Mar;15(3):1117-38. doi: 10.1074/mcp.M115.052498. Epub 2015 Dec 28. Mol Cell Proteomics. 2016. PMID: 26711468 Free PMC article.
-
Caldesmon and the regulation of cytoskeletal functions.Adv Exp Med Biol. 2008;644:250-72. doi: 10.1007/978-0-387-85766-4_19. Adv Exp Med Biol. 2008. PMID: 19209827 Free PMC article. Review.
-
Myosin IIB deficiency in embryonic fibroblasts affects regulators and core members of the par polarity complex.Histochem Cell Biol. 2011 Sep;136(3):245-66. doi: 10.1007/s00418-011-0840-0. Epub 2011 Jul 23. Histochem Cell Biol. 2011. PMID: 21785947
-
The actin-binding domain of cortactin is dynamic and unstructured and affects lateral and longitudinal contacts in F-actin.Cell Motil Cytoskeleton. 2009 Feb;66(2):90-8. doi: 10.1002/cm.20328. Cell Motil Cytoskeleton. 2009. PMID: 19089942 Free PMC article.
-
Differential effects of caldesmon on the intermediate conformational states of polymerizing actin.J Biol Chem. 2010 Jan 1;285(1):71-9. doi: 10.1074/jbc.M109.065078. Epub 2009 Nov 4. J Biol Chem. 2010. PMID: 19889635 Free PMC article.
References
-
- Bursac P, Lenormand G, Fabry B, Oliver M, Weitz DA, Viasnoff V, Butler JP, Fredberg JJ. Cytoskeletal remodelling and slow dynamics in the living cell. Nat Mater. 2005;4(7):557–61. - PubMed
-
- Clarkson MR, Murphy M, Gupta S, Lambe T, Mackenzie HS, Godson C, Martin F, Brady HR. High glucose-altered gene expression in mesangial cells. Actin-regulatory protein gene expression is triggered by oxidative stress and cytoskeletal disassembly. J Biol Chem. 2002;277(12):9707–12. - PubMed
-
- Cortes P, Mendez M, Riser BL, Guerin CJ, Rodriguez-Barbero A, Hassett C, Yee J. F-actin fiber distribution in glomerular cells: structural and functional implications. Kidney Int. 2000;58(6):2452–61. - PubMed
-
- D’Angelo G, Graceffa P, Wang CLA, Wrangle J, Adam LP. Mammal-specific, ERK-dependent, caldesmon phosphorylation in smooth muscle. Quantitation using novel anti-phosphopeptide antibodies. J Biol Chem. 1999;274(42):30115–21. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
