

Abrogation of the CLK-2 checkpoint leads to tolerance to base-excision repair intermediates
- PMID: 16964178
- PMCID: PMC1618380
- DOI: 10.1038/sj.embor.7400782
Abrogation of the CLK-2 checkpoint leads to tolerance to base-excision repair intermediates
Abstract
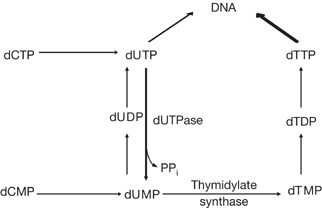
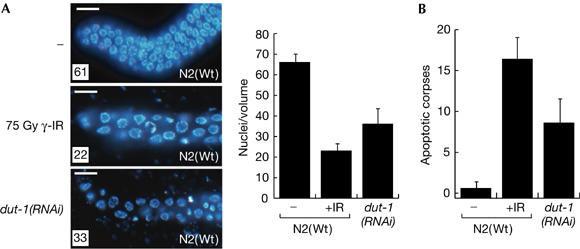
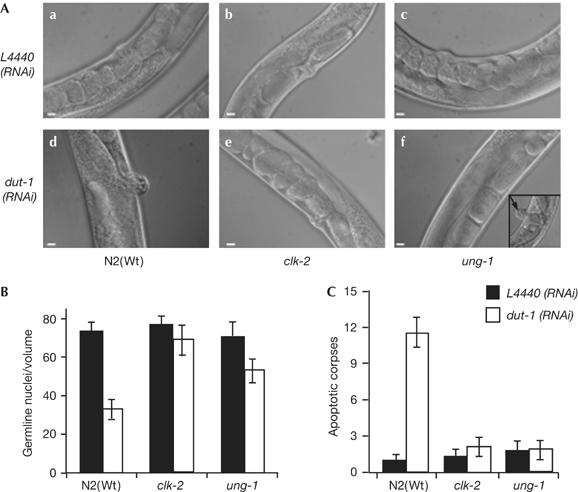
Incorporation of uracil during DNA synthesis is among the most common types of endogenously generated DNA damage. Depletion of Caenorhabditis elegans dUTPase by RNA interference allowed us to study the role of DNA damage response (DDR) pathways when responding to high levels of uracil in DNA. dUTPase depletion compromised development, caused embryonic lethality and led to activation of cell-cycle arrest and apoptosis. These phenotypes manifested as a result of processing misincorporated uracil by the uracil-DNA glycosylase UNG-1. Strikingly, abrogation of the clk-2 checkpoint gene rescued lethality and developmental defects, and eliminated cell-cycle arrest and apoptosis after dUTPase depletion. These data show a genetic interaction between UNG-1 and activation of the CLK-2 DDR pathway after uracil incorporation into DNA. Our results indicate that persistent repair intermediates and/or single-stranded DNA formed during repair of misincorporated uracil are tolerated in the absence of the CLK-2 checkpoint in C. elegans.
Figures

References
-
- Ahmed S, Alpi A, Hengartner MO, Gartner A (2001) C. elegans RAD-5/CLK-2 defines a new DNA damage checkpoint protein. Curr Biol 11: 1934–1944 - PubMed
-
- Alpi A, Pasierbek P, Gartner A, Loidl J (2003) Genetic and cytological characterization of the recombination protein RAD-51 in Caenorhabditis elegans. Chromosoma 112: 6–16 - PubMed
-
- Barnes DE, Lindahl T (2004) Repair and genetic consequences of endogenous DNA base damage in mammalian cells. Annu Rev Genet 38: 445–476 - PubMed
-
- Bartkova J et al. (2005) DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature 434: 864–870 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
