

A genomic view of the sea urchin nervous system
- PMID: 16965768
- PMCID: PMC1950334
- DOI: 10.1016/j.ydbio.2006.08.007
A genomic view of the sea urchin nervous system
Abstract
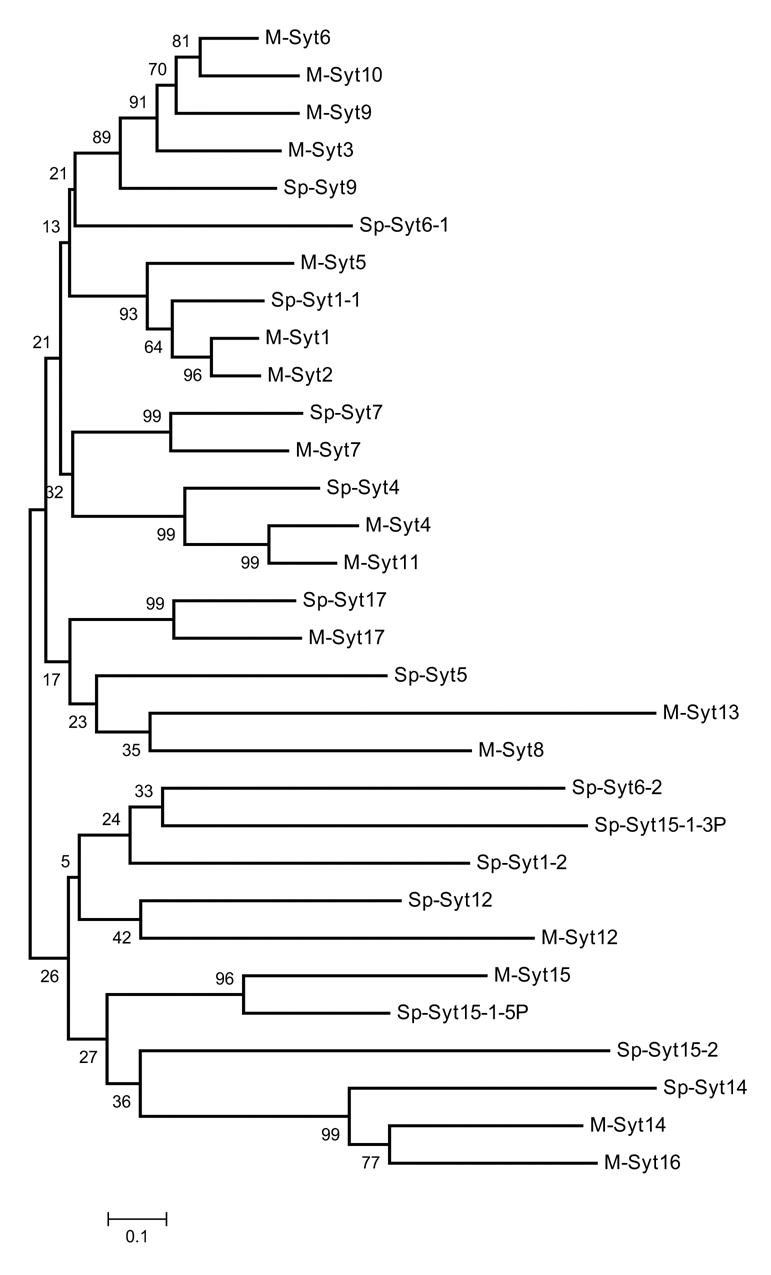
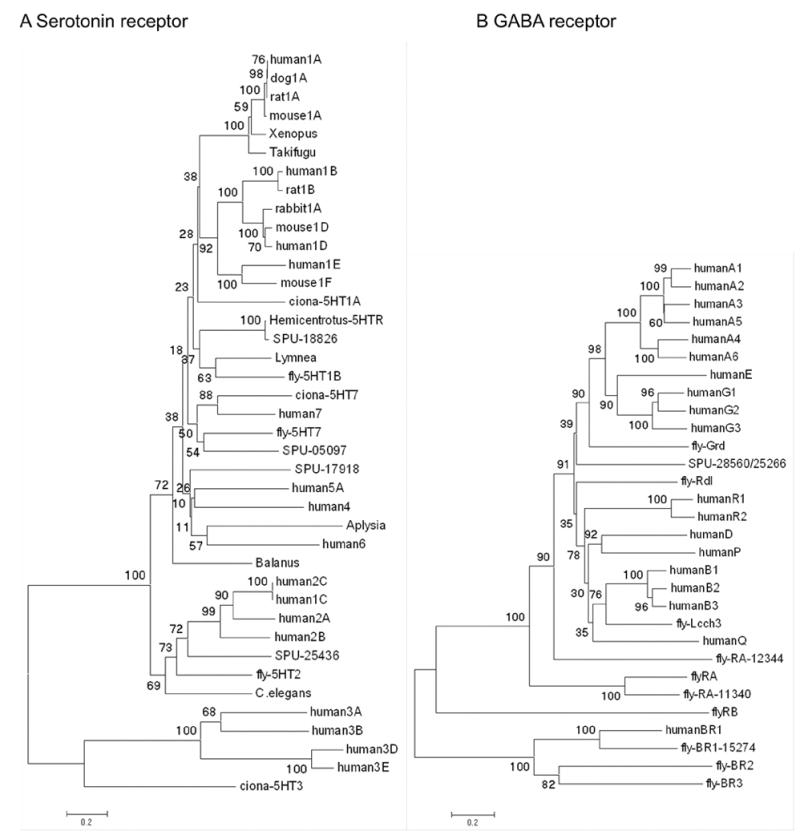
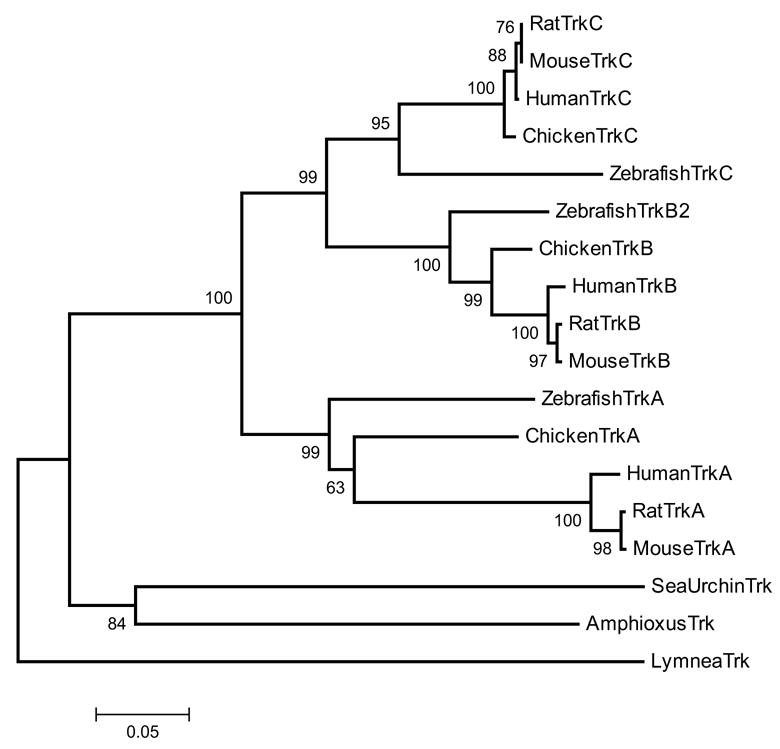
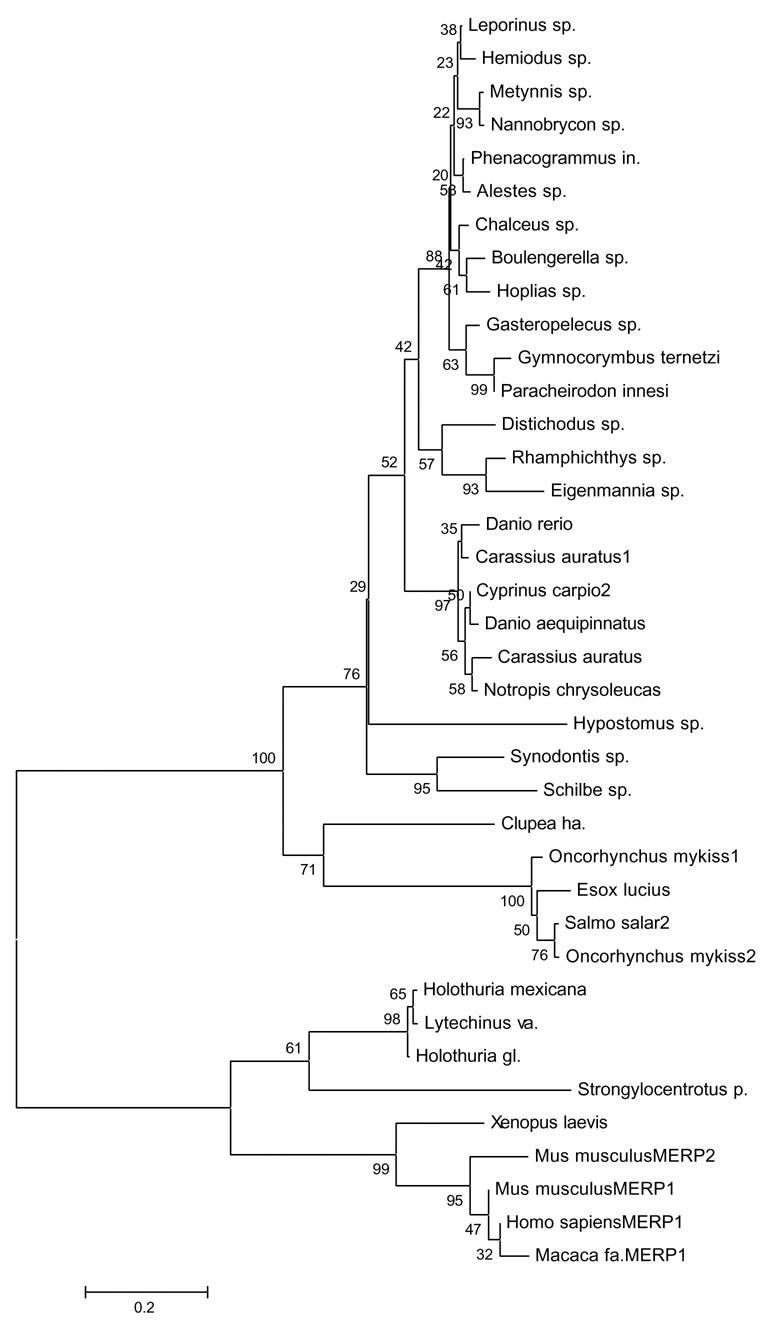
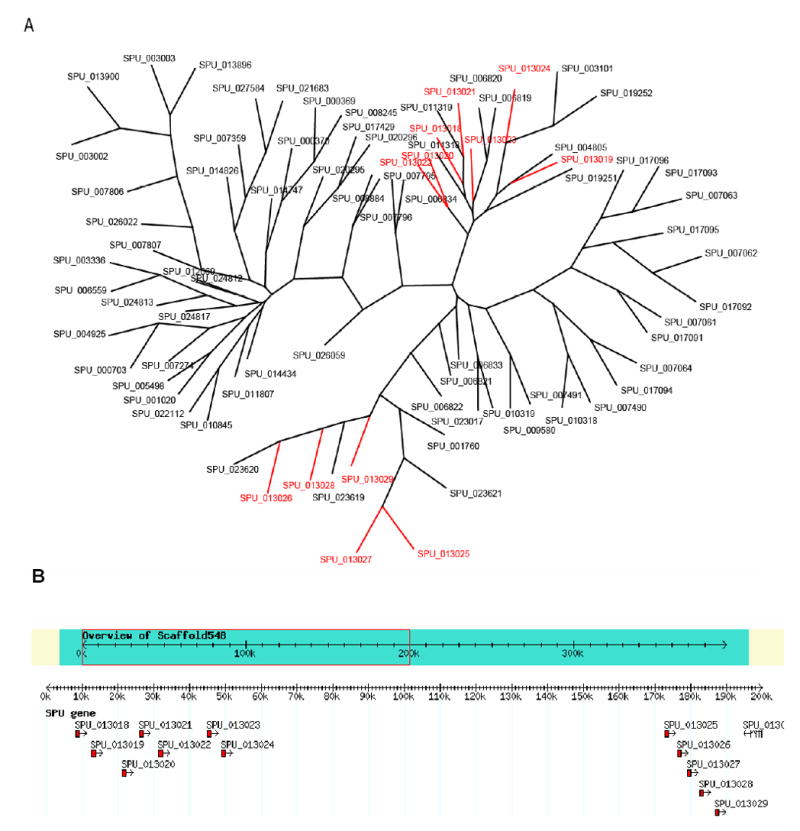
The sequencing of the Strongylocentrotus purpuratus genome provides a unique opportunity to investigate the function and evolution of neural genes. The neurobiology of sea urchins is of particular interest because they have a close phylogenetic relationship with chordates, yet a distinctive pentaradiate body plan and unusual neural organization. Orthologues of transcription factors that regulate neurogenesis in other animals have been identified and several are expressed in neurogenic domains before gastrulation indicating that they may operate near the top of a conserved neural gene regulatory network. A family of genes encoding voltage-gated ion channels is present but, surprisingly, genes encoding gap junction proteins (connexins and pannexins) appear to be absent. Genes required for synapse formation and function have been identified and genes for synthesis and transport of neurotransmitters are present. There is a large family of G-protein-coupled receptors, including 874 rhodopsin-type receptors, 28 metabotropic glutamate-like receptors and a remarkably expanded group of 161 secretin receptor-like proteins. Absence of cannabinoid, lysophospholipid and melanocortin receptors indicates that this group may be unique to chordates. There are at least 37 putative G-protein-coupled peptide receptors and precursors for several neuropeptides and peptide hormones have been identified, including SALMFamides, NGFFFamide, a vasotocin-like peptide, glycoprotein hormones and insulin/insulin-like growth factors. Identification of a neurotrophin-like gene and Trk receptor in sea urchin indicates that this neural signaling system is not unique to chordates. Several hundred chemoreceptor genes have been predicted using several approaches, a number similar to that for other animals. Intriguingly, genes encoding homologues of rhodopsin, Pax6 and several other key mammalian retinal transcription factors are expressed in tube feet, suggesting tube feet function as photosensory organs. Analysis of the sea urchin genome presents a unique perspective on the evolutionary history of deuterostome nervous systems and reveals new approaches to investigate the development and neurobiology of sea urchins.
Figures


Similar articles
-
Generation, annotation, evolutionary analysis, and database integration of 20,000 unique sea urchin EST clusters.Genome Res. 2003 Dec;13(12):2736-46. doi: 10.1101/gr.1674103. Genome Res. 2003. PMID: 14656975 Free PMC article.
-
Evolutionary convergence in Otx expression in the pentameral adult rudiment in direct-developing sea urchins.Dev Genes Evol. 2003 Mar;213(2):73-82. doi: 10.1007/s00427-003-0299-7. Epub 2003 Feb 8. Dev Genes Evol. 2003. PMID: 12632176
-
NGFFFamide and echinotocin: structurally unrelated myoactive neuropeptides derived from neurophysin-containing precursors in sea urchins.J Exp Biol. 2009 Apr;212(Pt 8):1067-77. doi: 10.1242/jeb.027599. J Exp Biol. 2009. PMID: 19329739
-
Identification and developmental expression of the ets gene family in the sea urchin (Strongylocentrotus purpuratus).Dev Biol. 2006 Dec 1;300(1):35-48. doi: 10.1016/j.ydbio.2006.08.012. Epub 2006 Aug 10. Dev Biol. 2006. PMID: 16997294 Review.
-
High regulatory gene use in sea urchin embryogenesis: Implications for bilaterian development and evolution.Dev Biol. 2006 Dec 1;300(1):27-34. doi: 10.1016/j.ydbio.2006.10.016. Epub 2006 Oct 18. Dev Biol. 2006. PMID: 17101125 Free PMC article. Review.
Cited by
-
Properties of cannabinoid-dependent long-term depression in the leech.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2010 Nov;196(11):841-51. doi: 10.1007/s00359-010-0566-9. Epub 2010 Aug 28. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2010. PMID: 20803022
-
The genome sequence of the protostome Daphnia pulex encodes respective orthologues of a neurotrophin, a Trk and a p75NTR: evolution of neurotrophin signaling components and related proteins in the bilateria.BMC Evol Biol. 2009 Oct 6;9:243. doi: 10.1186/1471-2148-9-243. BMC Evol Biol. 2009. PMID: 19807921 Free PMC article.
-
Spatial and temporal patterns of gene expression during neurogenesis in the sea urchin Lytechinus variegatus.Evodevo. 2019 Feb 12;10:2. doi: 10.1186/s13227-019-0115-8. eCollection 2019. Evodevo. 2019. PMID: 30792836 Free PMC article.
-
Rx and its downstream factor, Musashi1, is required for establishment of the apical organ in sea urchin larvae.Front Cell Dev Biol. 2023 Aug 15;11:1240767. doi: 10.3389/fcell.2023.1240767. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37655161 Free PMC article.
-
Biochemical, Anatomical, and Pharmacological Characterization of Calcitonin-Type Neuropeptides in Starfish: Discovery of an Ancient Role as Muscle Relaxants.Front Neurosci. 2018 Jun 8;12:382. doi: 10.3389/fnins.2018.00382. eCollection 2018. Front Neurosci. 2018. PMID: 29937709 Free PMC article.
References
-
- Adams D, Hasson B, Boyer-Boiteau A, El-Khishin A, Shashoua V. A peptide fragment of ependymin neurotrophic factor uses protein kinase C and the mitogen-activated protein kinase pathway to activate c-Jun N-terminal kinase and a functional AP-1 containing c-Jun and c-Fos proteins in mouse NB2a cells. J Neurosci Res. 2003;72:405–416. - PubMed
-
- Adayev T, Ranasinghe B, Banerjee P. Transmembrane signaling in the brain by serotonin, a key regulator of physiology and emotion. Bioscience Reports. 2005;25:363–385. - PubMed
-
- Aizenberg J, Tkachenko A, Weiner S, Addadi L, Hendler G. Calcitic microlenses as part of the photoreceptor system in brittlestars. Nature. 2001;412:819–822. - PubMed
-
- Arac D, Chen X, Khant HA, Ubach J, Ludtke SJ, Kikkawa M, Johnson AE, Chiu W, Sudhof TC, Rizo J. Close membrane-membrane proximity induced by Ca(2+)-dependent multivalent binding of synaptotagmin-1 to phospholipids. Nat Struct Mol Biol. 2006;13:209–17. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
